 2021, Vol. 48
2021, Vol. 48文章信息
引用本文 |

基金项目
- 国家自然科学基金(41701341);广东省自然科学基金面上项目(2019A1515011737);广东省现代农业产业技术体系项目(2019KJ140);广东省农业科学院科技创新战略专项资金(高水平农科院建设)-人才项目(R2017PY-QF005);广东省农业科学院科技创新战略专项资金(高水平农科院建设)-“十三五”学科团队建设项目(201623TD)
作者简介
- 涂玉婷(1987-),女,博士,副研究员,研究方向为农业资源利用与新型农业功能材料研发,E-mail:sysutyt@hotmail.com.
通讯作者
- 彭智平(1964-),男,研究员,研究方向为新型肥料和土壤调理剂开发,E-mail:ytifei@aliyun.com.
文章历史
- 收稿日期:2020-12-11

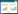


【研究意义】化感自毒效应是植物通过地上部及其凋落物的挥发、淋溶以及根系分泌和植株残茬腐解等途径,释放出的次生代谢物,对自身或种内其它植物生长产生危害的一种现象[1]。已有研究表明,高等植物分泌的化感物质主要是酚酸类和萜类,以及少数含氮化合物、聚乙炔和香豆素等次生物质。其中酚酸类化感物质因其活性强,相对总量高,被研究者们作为化感自毒物质的重点研究对象[2-3]。高浓度的酚酸类化感物质通过影响植物细胞膜透性、水分和养分吸收、光合速率、抗氧化酶活性等途径对植物的生长代谢过程和生长调节系统产生抑制作用;自毒物质对根际微生物具有趋化作用,使有益微生物减少,病原微生物增加[4-5]。近年来快速发展的设施蔬菜栽培技术存在种植结构单一、复种指数高的特点,这种连作模式下,土壤中酚酸类物质的累积导致的自毒作用和土壤微生态失衡,已被证实是造成连作障碍的根本原因[6]。开发去除连作土壤中化感自毒物质的技术,并明确其作用机制,是从根源上解决连作障碍的有效措施。
【前人研究进展】为了缓解日益严重的连作障碍问题,实际生产中采用客土法、土壤熏蒸、施用微生物制剂、施用土壤改良剂等,其中施用生物炭已经被认为是一种成本较低、效果稳定的方法之一[7]。生物炭是以生物质为原料,在高温缺氧条件下热解产生的一类具有微孔结构丰富、比表面积大、稳定性强等一系列优异性质的多功能材料。武春成等[8]研究发现按连作土壤质量比5% 施入玉米秸秆生物炭,可使土壤pH提高0.56个单位,速效钾含量提高8.6%,有效降低土壤真菌和尖孢镰刀菌数量,提高细菌数量,细菌/真菌比值是对照的14.3倍,通过对黄瓜根区土壤微生态环境的改善,促进了黄瓜生长和产量的提高。已有研究表明,生物炭的施用能有效降低土壤中残余酚酸含量,并一定程度地缓解连作条件下化感物质对番茄、黄瓜、苹果、烤烟等作物的化感胁迫效应[9-12],还具有改善连作土壤理化性质、提高土壤酶活性、优化微生物群落结构的作用[13-15]。
【本研究切入点】目前已有的研究大多关注外源生物炭对化感胁迫下“植株-土壤-微生物”体系中某一个或几个元素的变化情况,缺乏对体系中各元素之间变化的关联性分析。番茄是世界上也是我国栽培面积最大的茄果类蔬菜之一,在连作条件下番茄根系分泌的酚酸类化感自毒物质极易在土壤中累积,从而引发较为严重的连作障碍问题。本研究以番茄为供试作物,通过盆栽试验,考察施用生物质炭对酚酸胁迫下番茄植株生长和生理指标、果实产量和品质、土壤理化性质及酚酸含量的影响,利用高通量测序技术研究番茄根际土壤中细菌和真菌群落结构对酚酸类化感物质和生物炭输入的响应情况,通过相关性分析和冗余分析揭示“生物炭-土壤环境因子-根际土壤微生物-植株”之间的关联性与相互作用机制。研究结果将为设施蔬菜产业的可持续发展提供理论依据和实践指导。【拟解决的关键问题】探索酚酸胁迫下“植株-土壤-微生物”体系中各因子对外源生物炭输入的响应特征及各因子之间变化的关联互作机制,对深入解析生物炭连作土壤生态调控效应,开发稳定高效的生物炭基连作土壤调理剂具有重要意义。
1 材料与方法 1.1 试验材料供试番茄品种千禧圣女果为广东地区栽培的主要小番茄品种之一。苯甲酸、香草酸、对羟基苯甲酸、阿魏酸、肉桂酸均购买自阿拉丁试剂(上海)有限公司。生物炭由花生壳在700 ℃下隔绝空气热解2 h制备,花生壳生物炭的产率为24.1%,灰分含量为1.4%,比表面积为83.4 m2/g,pHPZC为8.33,阳离子交换量为7.42 cmol/kg。供试土壤为赤红壤,前茬为水稻。土壤pH 6.21,EC值0.24 μs/cm,有机质2.15%,碱解氮93.54 mg/kg,速效磷81.82 mg/kg,速效钾59.8 mg/kg,全氮1.01 g/kg,全磷1.10 g/kg,全钾12.25 g/kg。
1.2 试验方法试验于2019年10月至2020年4月在广东省农业科学院农业资源与环境研究所温网室(113°20′54.63″E,23°08′47.56″N)进行。试验用直径26.0 cm、高24.0 cm的塑料盆,每盆装入已去除石砾和动植物残体的风干土壤6.5 kg。试验设3个处理:CK,风干土壤+1.3 L无菌水(含水率20%);T1处理,风干土壤+1.3 L混合酚酸溶液;T2处理,风干土壤+5% 生物炭+1.3 L混合酚酸溶液。试验参照张玥琦等[16]和李亮亮等[17]研究确定的番茄根系和根际土壤中酚酸的种类和浓度进行混合酚酸溶液的配制,施用后,使土壤中酚酸类物质的初始浓度为苯甲酸50 μg/g、香草酸40 μg/g、对羟基苯甲酸25 μg/g、阿魏酸10 μg/g、肉桂酸10 μg/g,总酚酸含量为135 μg/g。挑选长至六叶一心且长势基本一致的番茄幼苗进行盆栽,每钵1株,每个处理3次重复,每个重复30株,此后每3 d浇1次水,以浇透而不渗出为度。采用单杆整枝,主枝长至2 m时打顶,每穗留6个果实。试验期间各处理采用一致的施肥管理。
1.3 测定项目及方法移栽后30 d和120 d,用百分之一电子天平分别对各处理番茄植株的生物量、地上部和地下部鲜重进行测定,采用TTC法(氯化三苯基四氮唑法)测定番茄根系活力。于2020年2月3日开始采收番茄果实,每次采收时各株单独计产,各次采摘的产量相加即为单株总产量,用万分之一电子天平称量并计算各处理平均单果重。统一选取各处理第2穗和第3穗小番茄进行果实品质测定,其中维生素C含量测定采用2, 6-二氯腚酚测定法,可溶性糖含量采用蒽酮比色法测定,有机酸含量采用酸碱滴定法测定。盛果期时每个处理选取5颗番茄采用抖土法收集各处理根际土壤,并均匀混合,一部分于-80 ℃超低温冰箱保存用于土壤残余酚酸含量和土壤微生物多样性的测定,另一部分风干后过1 mm筛用于土壤常规理化指标测定。
土壤pH值采用水土比2.5 ∶ 1 pH计法测定;土壤电导率采用电导率仪法测定;土壤有机质采用重铬酸钾外加热法测定;碱解氮用碱解扩散硼酸吸收法;速效磷采用钼锑抗比色法测定;速效钾采用NH4OAc浸提-火焰光度法测定;土壤全氮采用全自动凯氏定氮仪测定;全磷采用NaOH熔融-钼锑抗比色法测定;全钾采用NaOH熔融-火焰光度法测定。
土壤中残余酚酸的提取参考李培栋等[18]的方法。使用1 mol/L NaOH,按土水比1 ∶ 1对土壤中的酚酸进行浸提24 h,并通过超声波处理30 min以提高提取效率,离心分离后用12 mol/L盐酸将溶液pH调整至2.5以沉淀胡敏酸。静置2 h后8 000 r/min离心10 min,上清液过0.22 μm滤膜后于4 ℃保存。滤液中总酚酸的测定采用福林酚比色法[19],滤液中苯甲酸、香草酸、对羟基苯甲酸、阿魏酸、肉桂酸的浓度采用Waters e2695 Alliance高效液相色谱检测,色谱柱为Water Symmetry C18(5.0 μm, 4.6 mm×250 mm),进样量为20 μL,柱温30 ℃,检测波长280 nm,流动相为0.1% 甲酸溶液/甲醇(70 ∶ 30, V/V),流速0.8 mL/min,采用峰面积外标法进行定量分析。
土壤细菌和真菌的微生物多样性测定由北京百迈客生物科技有限公司完成。采用德国Macherey-Nagel公司的NucleoSpin 96 Soi土壤DNA提取试剂盒提取土壤总DNA,按照试剂盒说明书进行操作,采用1.5%琼脂糖凝胶电泳检测提取DNA的纯度和浓度,检测合格后对土壤DNA进行PCR扩增和高通量测序。土壤细菌16S rRNA(V3+V4)区的上游扩增引物为338F(5'-ACTCCTACGGGAGGCAGCA-3');下游引物为806R(5'-GGACTACHVGGGTWTCTAAT-3')。真菌I T S 1区上下游扩增引物分别为I T S 1 F(5'-CTTGGTCATTTAGAGGAAGTAA-3')和ITS2(5'-GCATCGATGAAGAACGCAGC-3')。在Illumina HiSeq平台上完成测序后,使用百迈客云平台进行细菌/真菌丰富度和多样性分析、菌群的分类学组成分析。
试验数据采用Microsoft Excel 2007和SPSS 19.0软件进行处理和统计分析,使用邓肯氏新复极差法进行差异显著性测验,使用Origin 9.0作图。
2 结果与分析 2.1 不同处理对番茄植株生长和生理指标的影响由图 1可知,施用外源酚酸的T1处理与CK相比,移栽后30 d、120 d番茄植株生物量分别减少23.6% 和10.0%,地下部鲜重分别减少37.2% 和24.0%,根冠比分别降低12.6% 和18.8%,根系活力分别下降39.2% 和48.5%,说明试验所添加的外源酚酸对番茄植株的根系活力、地下部和地上部的生长均表现出显著抑制作用;施用生物炭的T2处理移栽后30 d、120 d番茄植株生物量比T1处理分别增加11.3% 和8.0%,地下部鲜重增加27.0% 和18.2%,根冠比分别提高15.2% 和6.3%,根系活力分别增加30.6% 和25.2%。与CK相比,移栽后30 d番茄苗各生长和生理指标的相对增幅均高于收获期。可见,施加适量的生物炭,有利于缓解酚酸对番茄幼苗根部和地上部的抑制效应,提高根系活力,改善植株形态特征,且这种调控作用在番茄苗期的改善效果要优于收获期。

|
| 图 1 不同处理番茄苗移栽后30 d、120 d的生物量 Fig. 1 Effects of different treatments on biomass of tomato plants at 30th and 120th day after transplanting A:生物量;B:地下部鲜重;C:根冠比;D:根系活力 A: Biomass; B: Root fresh weight; C: Root-shoot ratio; D: Root activity |
2.2 不同处理对番茄果实产量和品质的影响
由表 1可知,与CK相比,施用外源酚酸后番茄单株产量和单果重分别降低9.5% 和11.9%,且差异显著。与T1处理相比,施用生物炭(T2处理)可显著提高酚酸胁迫下番茄单株产量和单果重,增幅分别为15.2% 和4.3%。番茄品质方面,外源酚酸的添加导致番茄维生素C和可溶性糖含量分别降低18.2% 和11.0%,有机酸含量增加26.2%,糖酸比降低29.9%。而添加生物炭能使酚酸胁迫下番茄果实的维生素C、可溶性糖和糖酸比分别增加17.6%、6.6% 和32.0%,有效提高番茄果实品质,改善果实风味。
2.3 不同处理对土壤理化性质的影响
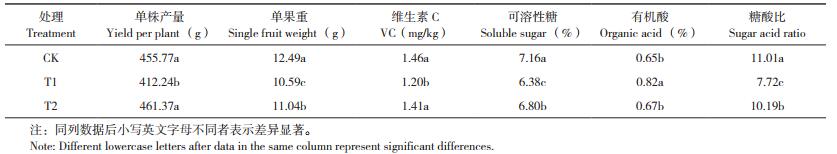
由表 2可知,与对照相比,施用外源酚酸处理的土壤pH值降低,有机质、碱解氮、有效钾、全氮、全钾含量均发生下降现象,降幅为2.1%~18.3%;土壤EC值、有效磷和全磷含量出现不同程度升高。施用生物炭可显著提高土壤pH值,降低EC值,并缓解外源酚酸对土壤养分指标的影响。与T1处理相比,T2处理土壤有机质、碱解氮、有效钾、全氮、全钾含量分别提高64.2%、6.1%、8.6%、13.8% 和5.8%。
|
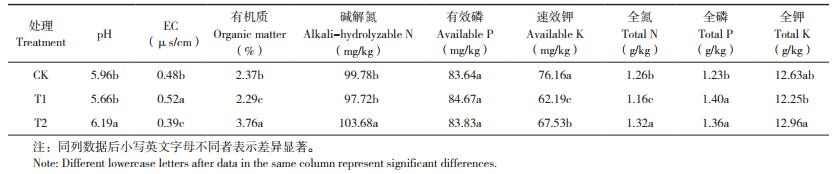
由表 3可知,T1处理土壤中残余的苯甲酸、香草酸、对羟基苯甲酸、阿魏酸、肉桂酸和总酚酸含量均显著高于CK。而与T1处理相比,T2处理可使土壤中5种残余酚酸含量降低14.6%~36.6%,总酚酸含量降低25.3%,表明生物炭能一定程度地降低土壤中残余酚酸含量。但是施用外源酚酸的T1处理土壤残余总酚酸含量仅比CK增加1.2 μg/g,占初始添加总酚酸用量的0.9%,说明施入土壤中的酚酸会经过土壤吸附、微生物分解等作用导致游离酚酸含量的降低。
|
2.4 不同处理对根际土壤微生物多样性的影响
由表 4可知,T1处理反映土壤样本细菌群落丰富度的ACE指数和Chao1指数分别比CK降低4.7% 和5.5%,且差异显著;辛普森指数和香农指数则可同时反映物种丰富度和均匀度,T1处理辛普森指数低于CK,香农指数比CK高5.9%。
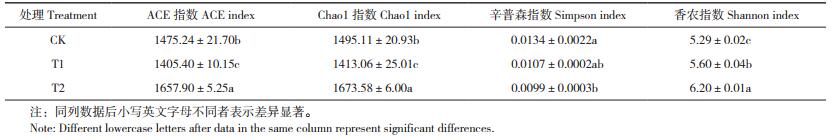
|
由表 5可知,与CK相比,T1处理反映土壤真菌丰富度的ACE指数和Chao1指数平均值分别增加34.1和32.4%,辛普森指数下降0.2%,香农指数增加6.7%,表明施用外源酚酸可导致番茄根际土壤细菌群落丰富度显著降低,真菌群落丰富度显著提升,而土壤中细菌和真菌群落的均匀度都得到一定程度的增加。与T1处理相比,施用生物炭(T2处理)可有效提升酚酸胁迫下土壤样品细菌群落ACE指数、Chao1指数和香农指数,分别提高18.0%、18.4% 和10.7%;真菌多样性指数则呈现相反的变化趋势,T2处理的土壤样品真菌群落ACE指数、Chao1指数和香农指数比T1处理分别降低10.3%、9.6% 和5.5%,且差异显著。由此可见,施用生物炭可显著增加酚酸胁迫下番茄根际土壤的细菌群落多样性,降低真菌群落多样性。
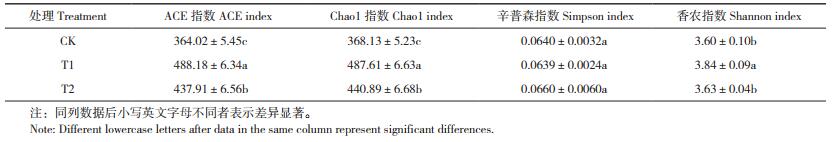
|
2.5 番茄产量、品质与土壤环境因子及微生物的相关性分析
对番茄单株产量和反应品质的糖酸比与土壤理化性质进行相关性分析,结果(表 6)表明,番茄产量与土壤pH、总氮的质量分数呈极显著正相关,与碱解氮和有效钾质量分数呈显著正相关,与残余总酚酸呈极显著负相关,与土壤有效磷质量分数呈显著负相关;糖酸比与有效钾、总氮质量分数呈极显著正相关,与pH也存在显著正相关关系,与有效磷、总磷、残余总酚酸质量分数呈显著负相关。
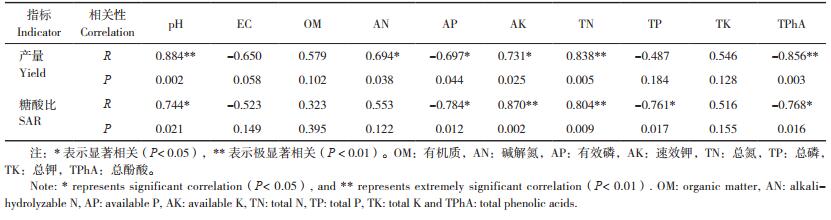
|
进一步选取与产量和品质有显著相关性的土壤环境因子,包括pH、碱解氮、有效磷、速效钾、总氮、总磷、总酚酸含量,采用冗余分析(Redundancy analysis, RDA)解析3个处理根际土壤细菌和真菌主要群落、土壤环境因子、番茄产量与品质之间的相关关系,结果(图 2)显示3个处理的细菌群落结构组成具有显著差异,RDA分析的轴1和轴2解释了土壤细菌纲水平上群落变异程度的74.0%。向量拟合结果显示番茄产量和品质与γ-变形菌纲(Gammaproteobacteria)、δ-变形菌纲(Deltaproteobacteria)、放线菌纲(Actinobacteria)的丰度呈正相关,与酸杆菌纲(Acidobacteriia)的丰度呈负相关。其中γ-变形菌纲、δ-变形菌纲的丰度与总氮、pH、碱解氮呈正相关,放线菌纲丰度与速效钾呈正相关。施用生物炭的T2处理与这3个土壤环境因子也呈正相关,说明施用外源生物炭可通过提高土壤pH、总氮、碱解氮含量,进而促进与番茄产量和品质呈正相关的变形菌丰度的增加。此外,施用生物炭处理还与芽孢杆菌纲(Gemmatimonadetes)丰度呈显著正相关。从图 2还可以看出,施用外源酚酸处理与土壤残余酚酸呈正相关,而TPhA与酸杆菌纲(Acidobacteriia)丰度呈正相关,表明施用外源酚酸会导致土壤总酚酸含量升高,从而促进与番茄产量和品质呈负相关的细菌群落丰度增加。

|
| 图 2 番茄产量、品质与土壤环境因子及细菌群落的冗余分析 Fig. 2 Redundancy analysis of tomato yield, quality and soil environmental factors and bacterial community SAR:番茄糖酸比,AN:土壤碱解氮,AP:有效磷,AK:速效钾,TN:总氮,TP:总磷,TPhA:总酚酸 SAR: tomato sugar acid ratio, AN: alkali-hydrolyzable N, AP: available P, AK: available K, TN: total N, TP: total P and TPhA: total phenolic acids |
3个处理根际土壤真菌群落结构与土壤环境因子和番茄产量、糖酸比的冗余分析结果(图 3)显示,前2个排序轴公共解释了不同处理真菌纲水平上群落结构变异的51.2%,属水平上群落丰度与番茄产量和品质呈正相关的真菌有伞菌属(Agaricomycetes)、小壶菌纲(Spizellomycetes),呈负相关的有座囊菌纲(Dothideomycetes)、粪壳菌纲(Sordariomycetes)、被孢霉纲(Mortierellomycetes)。T1处理与座囊菌纲(Dothideomycetes)、粪壳菌纲(Sordariomycetes)、被孢霉纲(Mortierellomycetes)的丰度呈正相关,T2处理则与伞菌纲(Agaricomycetes)、小壶菌纲(Spizellomycetes)丰度呈正相关。可见,外源酚酸的添加会导致与番茄产量和品质呈负相关的细菌和真菌群落丰度的增加,而添加生物炭有利于促进与番茄产量和品质呈正相关的细菌和真菌群落丰度的提高。

|
| 图 3 番茄产量、品质与土壤环境因子及真菌群落的冗余分析 Fig. 3 Redundancy analysis of tomato yield, quality and soil environmental factors and fungal community |
3 讨论
本试验所用酚酸浓度下,番茄根系活力出现显著下降,植株生物量、单株产量、单果重、可溶性糖、维生素C含量均出现明显下降,表明酚酸胁迫下番茄植株生长受到抑制,果实品质变劣。这一结果与已有的研究报道相似。翟彩霞等[20]研究发现每公斤土壤添加100~200 μg/mL苯甲酸或香豆素对番茄苗期的生长具有显著的抑制作用,具体表现为株高降低0.35~2.02 cm,复叶数、复叶长和叶面积减少;张恩平等[21]的研究表明,苯甲酸和肉桂酸的施用均会导致番茄幼苗根部丙二醛含量的增加,并打破番茄根部保护酶系统原有的平衡,造成根系的膜质过氧化。这主要是由于酚酸类化感物质可引发植物体内活性氧自由基的大量生成,同时,酚酸类化感物质对植株体内的抗氧化酶活性具有抑制作用,阻碍自由基的清除,过多的自由基易攻击细胞膜结构中的多不饱和脂肪酸,造成细胞膜结构氧化损伤,细胞膜透性升高,根系活力降低,严重影响根系对营养物质的吸收,从而导致植株生长受限、果实品质下降[2, 5]。施用生物炭可显著缓解酚酸对番茄生长的抑制作用。本研究中添加5% 生物炭可有效促进番茄根系发育,提高植株生物量和果实产量,果实的可溶性糖、维生素C含量和糖酸比得到提高,与酚酸胁迫处理相比,添加生物炭可有效改善番茄品质。金岩等[9]、吴珏等[22]的研究也发现添加生物炭对番茄具有促生、增产、提质的效果,与本研究结果一致。
为了探究生物炭对酚酸胁迫下番茄生长的促进和果实品质提升的作用机制,本研究进一步测定了3个处理中土壤理化性质和番茄根际土壤微生物群落结构变化情况。结果表明,外源酚酸的添加导致土壤EC值和磷含量增加,pH、有机质、氮和钾含量均出现不同程度的下降。这与陈龙池等[23]的研究结果相似。由于酚酸类物质可以与土壤中的Al、Fe、Mn、P结合形成复合物,导致可溶性盐类含量增加,EC值升高,同时增加土壤中磷的供给。此外,高浓度酚酸还会抑制与土壤碳循环相关的多酚氧化酶和蔗糖酶的活性,降低与氮素转化相关的脲酶和蛋白酶活性,从而导致土壤有机质含量下降,供氮能力减弱[24-25]。
添加生物炭的处理,可有效缓解酚酸对土壤理化性质的影响,表现为土壤pH、有机质、氮和钾含量显著提高。已有研究表明,由于生物炭具有较强的碱性,施入土壤后可显著提高土壤pH值。此外,生物炭具有丰富的孔隙结构,对土壤水分、盐分、养分吸附能力强,可有效降低土壤EC值,保持土壤肥力[26]。番茄收获期土壤残余酚酸的测定结果表明,添加生物炭可有效降低土壤中残余酚酸的含量,这种去除作用一方面源于生物炭对酚酸的吸附[27],另一方面可能源于生物炭输入导致土壤微生物群落结构发生变化,从而加速酚酸的生物降解。
本研究对根际土壤微生物α多样性分析结果表明,外源酚酸的施入总体上降低土壤细菌多样性,提高真菌多样性。已有研究表明,酚酸类化感物质能够选择性地抑制根际土壤中氨化细菌、硝化细菌、根际促生细菌,酸化的土壤环境对真菌的生长具有促进作用[28-29]。添加生物炭可促进酚酸胁迫下细菌微生物多样性的增加,并降低真菌多样性。这主要是由于施入生物炭引起土壤酸碱度、电导率、水分等理化指标发生改变,升高的pH值有利于抑制真菌的繁殖,碳氮比的优化则有利于促进细菌丰度的增加。本研究的相关性分析结果表明提高土壤pH、碱解氮、有效磷和总氮含量,降低有效磷、总磷和土壤总酚酸含量,均有利于番茄产量和品质的提升。冗余分析结果表明,以上土壤理化性质的改变有利于促进变形菌、放线菌、芽孢杆菌、小壶菌丰度的增加,研究已证实这些菌中具有较多根际促生菌和病原菌拮抗菌。酸杆菌常作为土壤恶化的生物学指标[30],冗余分析表明酸杆菌丰度与番茄产量和品质呈负相关。施用外源酚酸处理会促进酸杆菌丰度增加,而添加生物炭与酸杆菌丰度呈负相关,即具有抑制作用。
4 结论本研究采用盆栽试验,并结合土壤理化性质测定方法和基于16S rRNA/ITS1基因的Illumina HiSeq高通量测序技术,研究施用生物炭对酚酸胁迫下番茄生长和生理特性、土壤理化性质、根际土壤微生物群落结构的影响。结果表明,与外源酚酸胁迫处理相比,添加生物炭可显著提高番茄植株提高根系活力,增加生物量和单株果实产量,并改善果实品质。5% 生物炭的添加量可显著提高酚酸胁迫下土壤的pH值,降低EC值和总酚酸含量,土壤有机质、碱解氮、有效钾、全氮、全钾均有不同程度提高。相关性分析表明,土壤pH、碱解氮、速效钾、总氮含量与番茄果实产量或品质存在显著或极显著的正相关性,而有效磷、总磷、总酚酸含量与番茄果实产量或品质存在显著或极显著的负相关性。冗余分析结果表明,与番茄果实产量和糖酸比呈正相关的变形菌钢(Proteobacteria)、放线菌钢(Actinobacteria)、伞菌钢(Agaricomycetes)、小壶菌钢(Spizellomycetes)的丰度与土壤总氮、pH、碱解氮、速效钾也存在一定程度的正相关。与番茄果实产量和糖酸比呈负相关的酸杆菌钢(Acidobacteriia)、座囊菌钢(Dothideomycetes)、粪壳菌钢(Sordariomycetes)、被孢霉钢(Mortierellomycetes)的丰度与土壤总酚酸含量呈正相关。说明生物炭施入导致土壤pH、碱解氮、速效钾等理化性质的提高和总酚酸含量的降低,土壤环境因子的改变能够驱动与番茄果实产量和品质呈正相关的微生物种群丰度的增加,并对番茄果实产量和品质呈负相关的微生物种群起到抑制作用。生物炭通过对土壤理化因子和微生物群落结构的双重调控作用,从而缓解酚酸类化感物质对番茄植株生长的抑制,实现果实产量和品质的提升。
| [1] |
李庆凯, 刘苹, 赵海军, 宋效宗, 林海涛, 沈玉文, 李林, 万书波. 玉米根系分泌物对连作花生土壤酚酸类物质化感作用的影响[J]. 中国农业科技导报, 2020, 22(3): 119-130. DOI:10.13304/j.nykjdb.2019.0482 LI Q K, LIU P, ZHAO H J, SONG X Z, LIN H T, SHEN Y W, LI L, WAN S B. Effects of maize root exudates on allelopathy of phenolic acids in soil of continuous cropping peanut[J]. Journal of Agricultural Science and Technology, 2020, 22(3): 119-130. DOI:10.13304/j.nykjdb.2019.0482 |
| [2] |
WARDANI D K, DARMANTI S, BUDIHASTUTI R. Allelochemical effect of Ageratum conyzoides L. leaf extract on soybean[Glycine max (L.)Merr. cv Grobogan]growth[J]. Journal of Physics: Conference Series, 2018, 1025: 1-8. DOI:10.1088/1742-6596/1025/1/012044 |
| [3] |
袁婷婷, 董坤, 郭增, 董艳. 阿魏酸诱导蚕豆枯萎病发生及根系组织结构损伤的化感效应[J]. 植物营养与肥料学报, 2020, 26(5): 914-923. DOI:10.11674/zwyf.19388 YUAN T T, DONG K, GUO Z, DONG Y. Allelopathic effects of ferulic acid inducing fusarium wilt occurrence and abnormal root tissue structure of faba bean[J]. Plant Nutrition and Fertilizer Science, 2020, 26(5): 914-923. DOI:10.11674/zwyf.19388 |
| [4] |
田给林, 毕艳孟, 孙振钧, 张路生. 酚酸类物质在作物连作障碍中的化感效应及其调控研究进展[J]. 中国科技论文, 2016, 11(6): 699-705. DOI:10.3969/j.issn.2095-2783.2016.06.022 TIAN G L, BI Y M, SUN Z J, ZHANG L S. Progressin allelopathic effect and regulation of phenolic acids for continuous cropping obstacle system[J]. China Sciencepaper, 2016, 11(6): 699-705. DOI:10.3969/j.issn.2095-2783.2016.06.022 |
| [5] |
LU X F, ZHANG H, LYU S S, DU G D, WANG X Q, WU C H, LYU D G. Effects of exogenous phenolic acids on photosystem functions and photosynthetic electron transport rate in strawberry leaves[J]. Photosynthetica, 2018, 56(2): 616-622. DOI:10.1007/s11099-017-0702-7 |
| [6] |
陈玲, 董坤, 杨智仙, 董艳, 汤利, 郑毅. 连作障碍中化感自毒效应及间作缓解机理[J]. 中国农学通报, 2017, 33(8): 91-98. DOI:10.11924/j.issn.1000-6850.casb16050045 CHEN L, DONG K, YANG Z X, DONG Y, TANG L, ZHENG Y. Allelopathy autotoxcity effect of successive cropping obstacle and its alleviate mechanism by intercropping[J]. Chinese Agricultural Science Bulletin, 2017, 33(8): 91-98. DOI:10.11924/j.issn.1000-6850.casb16050045 |
| [7] |
ATUCHA A, LITUS G. Effect of biochar amendments on peach replant disease[J]. Hortscience, 2015, 50(6): 863-868. DOI:10.21273/HORTSCI.50.6.863 |
| [8] |
武春成, 王彩云, 曹霞, 张慎好, 罗晓飞, 姜涛. 不同用量生物炭对连作土壤改良及黄瓜生长的影响[J]. 北方园艺, 2017(19): 150-154. DOI:10.11937/bfyy.20170690 WU C C, WANG C Y, CAO X, ZHANG S H, LUO X F, JIANG T. Effects of different biochar application rate on improvement of continuous cropping soil and cucumber growth[J]. Northern Horticulture, 2017(19): 150-154. DOI:10.11937/bfyy.20170690 |
| [9] |
金岩, 杨凤军, 吴瑕, 宋雅超, 赵长江. 套作与生物炭互作提高番茄土壤酶活性和果实品质[J]. 新农业, 2018(7): 6-10. JIN Y, YANG F J, WU X, SONG Y C, ZHAO C J. Intercropping and biochar interaction to improve tomato soil enzyme activity and fruit quality[J]. New Agriculture, 2018(7): 6-10. |
| [10] |
WANG Y, PAN F, WANG G, ZHANG G, WANG Y, CHEN X, MAO Z. Effects of biochar on photosynthesis and antioxidative system of Malus hupehensis Rehd. seedlings under replant conditions[J]. Scientia Horticulturae, 2014, 175: 9-15. DOI:10.1016/j.scienta.2014.05.029 |
| [11] |
王雪玉, 马建华, 李明, 刘金泉, 胡云, 田勇. 生物炭对不同种植年限土壤黄瓜生长及真菌丰度的影响[J]. 中国瓜菜, 2018, 31(11): 22-25. DOI:10.3969/j.issn.1673-2871.2018.11.005 WANG X Y, MA J H, LI M, LIU J Q, HU Y, TIAN Y. Effects of biochar on cucumber growth and fungus abundance of soil in different planting years[J]. Chinese Melon, 2018, 31(11): 22-25. DOI:10.3969/j.issn.1673-2871.2018.11.005 |
| [12] |
刘卉, 荆永锋, 周清明, 黎娟, 向德明. 连续施用生物炭对连作烤烟烟叶主要化学成分的影响[J]. 西南农业学报, 2019, 32(1): 202-207. DOI:10.16213/j.cnki.scjas.2019.1.033 LIU H, JING Y F, ZHOU Q M, LI J, XIANG D M. Effects of continuous application of biochar on main chemical components in tobacco leaves of flue-cured tobacco continuous cropping[J]. Southwest China Journal of Agricultural Sciences, 2019, 32(1): 202-207. DOI:10.16213/j.cnki.scjas.2019.1.033 |
| [13] |
顾美英, 杨蓉, 徐万里, 唐光木, 张志东, 张云舒, 冯雷, 王宁, 詹发强. 棉秆炭配施生物有机肥对连作棉花根际土壤微生态和棉花生长的影响[J]. 中国农业科技导报, 2019, 21(10): 47-57. DOI:10.13304/j.nykjdb.2018.0755 GU M Y, YANG R, XU W L, TANG G M, ZHANG Z D, ZHANG Y S, FENG L, WANG N, ZHAN F Q. Effects of cotton stalk biochar combined with bio-organic fertilizer on rhizosphere soil micro-ecology and cotton growth of continuous cropping cotton[J]. Journal of Agricultural Science and Technology, 2019, 21(10): 47-57. DOI:10.13304/j.nykjdb.2018.0755 |
| [14] |
武春成, 李天来, 曹霞, 张勇勇, 杨丽娟. 添加生物炭对连作营养基质理化性质及黄瓜生长的影响[J]. 核农学报, 2014, 28(8): 1534-1539. DOI:10.11869/j.issn.100-8551.2014.08.1534 WU C C, LI T L, CAO X, ZHANG Y Y, YANG L J. Physicochemical properties of nutrition medium and cucumber growth under continuous cropping[J]. Journal of Nuclear Agricultural Sciences, 2014, 28(8): 1534-1539. DOI:10.11869/j.issn.100-8551.2014.08.1534 |
| [15] |
王彩云, 武春成, 曹霞, 贺字典, 曾晓玉, 姜涛. 生物炭对温室黄瓜不同连作年限土壤养分和微生物群落多样性的影响[J]. 应用生态学报, 2019, 30(4): 1359-1366. DOI:10.13287/j.1001-9332.201904.036 WANG C Y, WU C C, CAO X, HE Z D, ZENG X Y, JIANG T. Effects of biochar on soil nutrition and microbial community diversity under continuous cultivated cucumber soils in greenhouse[J]. Chinese Journal of Applied Ecology, 2019, 30(4): 1359-1366. DOI:10.13287/j.1001-9332.201904.036 |
| [16] |
张玥琦, 刘慧, 赵凤艳, 张国显, 李明静, 杨丽娟. 不同施肥措施对番茄连作土壤酚酸含量和微生物功能多样性的调节[J]. 土壤通报, 2017, 48(4): 887-894. DOI:10.19336/j.cnki.trtb.2017.04.17 ZHANG Y Q, LIU H, ZHAO F Y, ZHANG G X, LI M J, YANG L J. Regulation of various fertilization measures on soil microbial functional diversity and phenolic acid contents under tomato continuous cropping in greenhouse[J]. Chinese Journal of Soil Science, 2017, 48(4): 887-894. DOI:10.19336/j.cnki.trtb.2017.04.17 |
| [17] |
李亮亮, 吴正超, 陈彬, 刘文娥, 臧健. 生物炭对添加自毒物质土壤酶活性、微生物区系结构的影响[J]. 华北农学报, 2015, 30(4): 219-225. DOI:10.7668/hbnxb.2015.04.037 LI L L, WU Z C, CHEN B, LIU W E, ZANG J. The effect of biochar on soil enzyme activity and microbial community structure in the soil under autotoxicity stress[J]. Acta Agriculturae Boreali-Sinica, 2015, 30(4): 219-225. DOI:10.7668/hbnxb.2015.04.037 |
| [18] |
李培栋, 王兴祥, 李奕林, 王宏伟, 梁飞燕, 戴传超. 连作花生土壤中酚酸类物质的检测及其对花生的化感作用[J]. 生态学报, 2010, 30(8): 2128-2134. LI P D, WANG X X, LI Y L, WANG H W, LIANG F Y, DAI C C. The contents of phenolic acids in continuous cropping peanut and their allelopathy[J]. Acta Ecologica Sinica, 2010, 30(8): 2128-2134. |
| [19] |
田给林, 严婷婷, 毕艳孟, 孙振钧, 张潞生. 草莓连作土壤灭菌与施用有机肥对根际土壤酚酸及土壤酶活性的影响[J]. 园艺学报, 2015, 42(10): 2039-2048. DOI:10.16420/j.issn.0513-353x.2015-0192 TIAN G L, YAN T T, BI Y M, SUN Z J, ZHANG L S. Effects of continuous cropping soil sterilization and applying of different fertilizer on phenolic acids in rhizosphere soil of the strawberry pants and soil enzyme activities[J]. Acta Horticulturae Sinica, 2015, 42(10): 2039-2048. DOI:10.16420/j.issn.0513-353x.2015-0192 |
| [20] |
翟彩霞, 张彦才, 王占武, 李洪涛, 李巧云. 土壤生态调控剂缓解酚酸类物质对番茄苗期生长抑制作用的效果[J]. 华北农学报, 2006, 21(z2): 54-58. DOI:10.3321/j.issn:1000-7091.2006.z2.013 ZHAI C X, ZHANG Y C, WANG Z W, LI H T, LI Q Y. The relieving effects of soil ecological control agents to phenolic acids on restraining growth of tomato seedlings[J]. Acta Agriculturae Boreali-Sinica, 2006, 21(z2): 54-58. DOI:10.3321/j.issn:1000-7091.2006.z2.013 |
| [21] |
张恩平, 张文博, 张淑红, 李亮亮, 李天来. 苯甲酸和肉桂酸对番茄幼苗根部保护酶及膜质过氧化的影响[J]. 西北农业学报, 2010, 19(1): 186-190. DOI:10.3969/j.issn.1004-1389.2010.01.040 ZHANG E P, ZHANG W B, ZHANG S H, LI L L, LI T L. Effects of exogenic benzoic acid and cinnamic acid on the root oxidative damage of tomato seedlings[J]. Acta Agriculturae Boreali-Occidentalis Sinica, 2010, 19(1): 186-190. DOI:10.3969/j.issn.1004-1389.2010.01.040 |
| [22] |
吴珏, 李建勇, 郭欣欣. 生物炭对设施菜田土壤理化性状及番茄生长的影响[J]. 蔬菜, 2020(4): 25-30. WU J, LI J Y, GUO X X. Effects of biochar on soil properties and growth of facility tomatoes[J]. Vegetables, 2020(4): 25-30. |
| [23] |
陈龙池, 廖利平, 汪思龙, 黄志群. 外源毒素对林地土壤养分的影响[J]. 生态学杂志, 2002, 21(1): 19-22. DOI:10.3321/j.issn:1000-4890.2002.01.005 CHEN L C, LIAO L P, WANG S L, HUANG Z Q. Effect of exotic toxin on the nutrition of woodland soil[J]. Chinese Journal of Ecology, 2002, 21(1): 19-22. DOI:10.3321/j.issn:1000-4890.2002.01.005 |
| [24] |
马云华, 王秀峰, 魏珉, 亓延凤, 李天来. 黄瓜连作土壤酚酸类物质积累对土壤微生物和酶活性的影响[J]. 应用生态学报, 2005, 16(11): 2149-2153. DOI:10.3321/j.issn:1001-9332.2005.11.028 MA Y H, WANG X F, WEI M, QI Y F, LI T L. Accumulation of phenolic acids in continuously cropped cucumber soil and their effects on soil microbes and enzyme activities[J]. Chinese Journal of Applied Ecology, 2005, 16(11): 2149-2153. DOI:10.3321/j.issn:1001-9332.2005.11.028 |
| [25] |
MOHAMMADKHANI N, SERVATI M. Nutrient concentration in wheat and soil under allelopathy treatments[J]. Journal of Plant Research, 2017, 131(1): 143-155. DOI:10.1007/s10265-017-0981-x |
| [26] |
沈盟, 蒋芳玲, 王珊, 唐静, 吴震. 生物质炭施用量对土壤性状和番茄产质量的影响[J]. 土壤, 2017, 49(3): 534-542. DOI:10.13758/j.cnki.tr.2017.03.016.tr.2017.03.016 SHEN M, JIANG F L, WANG S, TANG J, WU Z. Effects of biochar application amount on soil characteristics, yield and fruit properties of tomato[J]. Soils, 2017, 49(3): 534-542. DOI:10.13758/j.cnki.tr.2017.03.016.tr.2017.03.016 |
| [27] |
王艳芳, 沈向, 陈学森, 吴树敏, 毛志泉. 生物炭对缓解对羟基苯甲酸伤害平邑甜茶幼苗的作用[J]. 中国农业科学, 2014, 47(5): 968-976. DOI:10.3864/j.issn.0578-1752.2014.05.013 WANG Y F, SHEN X, CHEN X S, WU S M, MAO Z Q. Effects of biochar on alleviation of the influence of p-hydroxybenzoic acid on physiological characteristics in malus hupehensis rehd. Seedlings[J]. Scientia Agricultura Sinica, 2014, 47(5): 968-976. DOI:10.3864/j.issn.0578-1752.2014.05.013 |
| [28] |
LI X, DING C, HUA K, ZHANG T, ZHANG Y, ZHAO L, YANG Y, LIU J, WANG X. Soil sickness of peanuts is attributable to modifications in soil microbes induced by peanut root exudates rather than to direct allelopathy[J]. Soil Biology and Biochemistry, 2014, 78(11): 149-159. DOI:10.1016/j.soilbio.2014.07.019 |
| [29] |
齐虹凌, 贺国强, 李恒全, 刘德育, 孙宏宇, 元野. 轮作与连作对烤烟不同生育期根际土壤细菌群落结构的影响[J]. 中国烟草学报, 2015, 21(5): 42-48. DOI:10.16472/j.chinatobacco.2015.103 QI H L, HE G Q, LI H Q, LIU D Y, SUN H Y, YUAN Y. Effects of rotational and continuous cropping on bacterial community structures in rhizospheric soil at different growth stages of flue-cured tobacco[J]. Acta Tabacaria Sinica, 2015, 21(5): 42-48. DOI:10.16472/j.chinatobacco.2015.103 |
| [30] |
HAN L, WANG Z, LI N, WANG Y, FENG J, ZHANG X. Bacillus amyloliquefaciens b1408 suppresses fusarium wilt in cucumber by regulating the rhizosphere microbial ommunity[J]. Applied Soil Ecology, 2019, 136: 55-56. DOI:10.1016/j.apsoil.2018.12.011 |