 2021, Vol. 48
2021, Vol. 48文章信息
引用本文 |

基金项目
- 山东省自然科学基金(ZR2020MC036);枣庄学院博士科研启动基金(2018BS032, 2018BS033)
作者简介
- 张治国(1978—),男,博士,讲师,研究方向为植物生理生态学,E-mail:nouszzg@126.com.
通讯作者
- 魏海霞(1981—),女,博士,副教授,研究方向为植物生理生态学,E-mail:weihaixia0721@163.com.
文章历史
- 收稿日期:2020-11-23



【研究意义】湿地是地球上水陆相互作用而形成的独特生态系统,具有丰富的生物多样性和最高的生态功能[1]。中国是世界上湿地资源最丰富的国家之一,总面积约3 848万hm2[2]。长期以来,由于人为活动和自然因素等影响,湿地面积减少萎缩,湿地生态系统功能退化[3]。植物叶功能性状与生态系统功能关系密切,越来越多的研究者将湿地植物叶功能性状的研究引入到湿地生态系统的保护和恢复工作中。湿地植物叶功能性状的测定不仅有助于理解和揭示湿地植物群落的构建机制、湿地生态系统对环境变化的响应机制和湿地生态系统碳循环,而且对湿地生态系统服务功能及服务价值的评估具有重要意义[4-7]。【前人研究进展】比叶面积(Specific leaf area, SLA)可表示为叶片光合面积与叶片干重之比,直接反映了植物捕获光照资源和碳获取的能力[8],是全球叶片经济型谱中最主要的叶功能性状[9]。叶干物质含量(Leaf dry matter content, LDMC)为叶片干重与叶片饱和鲜重的比值,是诸多植物叶功能性状中易于测定的一个指标,可指示植物对养分的保存能力[10]。SLA和LDMC之间普遍存在负相关关系,二者常一起作为植物资源利用分类轴上划分植物种类的最佳变量,可综合反映植物对环境资源的利用能力以及适应环境的生存对策[11-13]。研究表明,植物在受到环境因子胁迫时,通常具有较小的SLA和较高的LDMC,植物保持体内水分和养分等资源的能力较强,一般采用资源保守型利用策略[14-15]。目前,国内有关SLA和LDMC的研究较多,但主要集中于对木本植物的分析上[16-19],而对于草本植物的研究较少,尤其是对于湿地草本植物的研究更少。【本研究切入点】桑沟湾湿地公园地处山东省荣成市,是我国第一个获批建立的国家城市湿地公园,动植物资源丰富,具有巨大的环境调节功能、景观美化和生态效益[20]。但是,由于该湿地属于浅海河口湿地,生长于此的植物常年受到较强海风的影响。大风可通过改变植物周围的气场环境以及增加对植物的机械刺激影而响植物的生存和生长[21]。研究表明,大风胁迫下,植物叶片变小变厚,SLA显著降低[22-23]。因此,研究该湿地主要植物SLA和LDMC的种间变化及其与其他湿地植物的差异,不仅有助于阐明该湿地植物对生存环境的适应机制,同时也可为该地区湿地建设中植物的筛选提供一定依据。【拟解决的关键问题】本研究以胶东半岛桑沟湾湿地公园的常见植物为研究对象,分别测定其SLA和LDMC,并与我们前期对鲁西南微山湖红荷湿地的测定数据进行比较分析。研究探讨问题如下:(1)不同科植物间SLA和LDMC的差异;(2)不同生活型以及水生和陆生植物间SLA和LDMC的差异;(3)不同湿地类型间湿地植物SLA和LDMC的差异。
1 材料与方法 1.1 研究区概况本研究主要在胶东半岛桑沟湾国家城市湿地公园进行,该湿地位于山东省荣成市市区东南部,总面积约13.91 km2,属于暖温带海洋季风气候,具有明显的干、湿季,年平均气温11.8 ℃,极端最高气温35.8 ℃,极端最低气温-18.3 ℃;年平均降水量760 mm,降水主要集中在5—9月;年平均风速6.8 m/s,大于8级的风35.8次;年平均日照时数2 526 h,全年无霜期约214 d。桑沟湾湿地是崖头河、沽河和十里河汇合入海口形成的浅海河口湿地,湿地内植物约有43科75种,湿地植物面积占湿地总面积的56. 87 %,对湿地的生态保存以及湿地环境的维持具有重要意义[20]。
为了比较不同湿地类型间植物叶性状的差异,本研究同时在鲁西南微山湖红荷湿地选择了与桑沟湾湿地相同的物种进行了样品采集。红荷湿地属于淡水草本沼泽湿地生态系统,位于山东省滕州市滨湖镇西北部,属于温带大陆季风气候,年平均气温13.8 ℃,年平均降水量770 mm,年平均风速2.5 m/s,年平均日照时数为2 384 h,全年无霜期216 d。
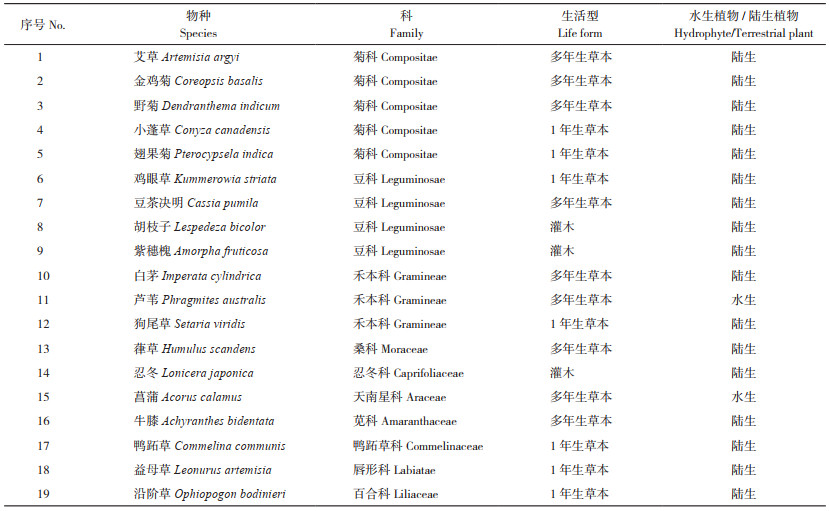
1.2 植物和土壤样品采集和测定在桑沟湾湿地,按不同生活型(灌木、多年生草本和一年生草本)采集常见19种植物的叶片,主要采样物种见表 1。19种植物大部分来自菊科(Compositae)、豆科(Leguminosae)和禾本科(Gramineae),其中芦苇和菖蒲为挺水植物,其他物种则为较耐湿的陆生植物(表 1)。
2018年7月,在设立的样地中,针对每种植物随机选取3株采集叶片,采集的叶片应完全展开且没有病虫害,之后将叶片装入密封袋中,带回室内分析。将叶片样品放入水中于5 ℃的黑暗环境中储藏12 h后取出,用吸水纸吸干叶片表面水分,之后在电子天平上称重(饱和鲜重)。然后用扫描仪(HP Scanjet2400)扫描叶片的单面面积,用Image Pro Plus 6.0软件对扫描图像进行处理获取叶片的实际叶面积。扫描后的叶片放入70 ℃烘箱中烘干48 h至恒重,之后取出称重。植物的SLA和LDMC分别采用下列公式计算:

|
土壤样品分别在样地的四角和中心位置进行采集。土壤样品通过土钻钻取获得,钻取深度为0~20 cm,之后将样品带回室内进行分析测定。将土壤样品自然风干碾碎后过孔径0.25 mm筛,然后采用凯式定氮法测定全氮[24]。
在微山湖红荷湿地,一共选择了与桑沟湾湿地相同的7个物种进行样品采集。7种植物分别为芦苇、菖蒲、葎草、艾草、白茅、小蓬草和紫穗槐。植物和土壤样品的采集和测定方法与桑沟湾湿地相同。
1.3 数据分析试验数据采用SPSS 16.0进行分析。用单因素方差分析和Tukey多重比较检验不同物种、不同生活型、不同类型湿地以及水生和陆生植物间SLA和LDMC的差异显著性。SLA和LDMC之间的相关关系用简单线性模型y = a + bx表示。
2 结果与分析 2.1 不同科植物间SLA和LDMC的差异从表 2可以看出,桑沟湾湿地19种植物的比叶面积(SLA)和叶干物质含量(LDMC)在不同植物间差异显著。其中,葎草和鸭跖草的SLA最大(315.8、308.1 cm2/g),而白茅和芦苇的SLA最小(134.6、130.3 cm2/g)。总的来说,SLA在19种植物间变异较大,变异系数为27.8%。LDMC在不同植物间变异也较大,19种植物的LDMC变化范围为0.150~0.458 g/g,变异系数为30.1%,其中LDMC较大的植物主要为芦苇(0.458 g/g)、白茅(0.430 g/g)、紫穗槐(0.426 g/g)和胡枝子(0.400 g/g),而较小的则为葎草(0.150 g/g)和鸭跖草(0.167 g/g)。

|
本研究中的19种植物分属10科,从SLA和LDMC在不同科植物间的变化情况(图 1)可以看出,SLA和LDMC在不同科植物间差异显著。在10科植物中,桑科和鸭跖草科植物的SLA最大(315.8±9.5、308.1±10.2 cm2/g),而LDMC最低(0.150±0.015、0.167±0.012 g/g);禾本科和豆科植物在10科植物中具有相对较高的LDMC(0.374±0.111、0.356±0.067 g/g)和较小的SLA(172.7±60.5、176.8± 28.3 cm2/g)。

|
| 图 1 不同科植物间SLA和LDMC比较 Fig. 1 Comparisons of SLA and LDMC among different plant family groups Mo:桑科;Cm:鸭跖草科;Am:苋科;La:唇形科;Co:菊科;Ca:忍冬科;Le:豆科;Gr:禾本科;Li:百合科;Ar:天南星科小写英文字母不同者表示不同科植物间差异显著 Mo: Moraceae; Cm: Commelinaceae; Am: Amaranthaceae; La; Labiatae; Co: Compositae; Ca: Caprifoliaceae; Le: Leguminosae; Gr: Gramineae; Li: Liliaceae; Ar: Araceae Different lowercase letters represent significant differences among different plant family groups |
2.2 不同生活型植物间SLA与LDMC的差异
本研究中的19种植物按照生活型可分为木本植物、多年生草本植物和1年生草本植物,从图 2可以看出,SLA和LDMC在3种生活型间差异显著。其中,SLA在3种生活型间从大到小依次为1年生草本植物(228.4±47.1 cm2/g)>多年生草本植物(198.3±63.5 cm2/g)>木本植物(163.7±20.4 cm2/g)(图 2A);LDMC在不同生活型间的大小顺序与SLA刚好相反,依次为木本植物(0.375±0.064 g/g)>多年生草本植物(0.308±0.095 g/g)>1年生草本植物(0.255±0.067 g/g)(图 2B)。

|
| 图 2 不同生活型植物间SLA和LDMC比较 Fig. 2 Comparisons of SLA and LDMC among different plant life forms Wo:木本植物Woody plant,Ph:多年生草本Perennial herd,Ah:1年生草本 Annual herd小写英文字母不同者表示不同生活型间差异显著 Different lowercase letters represent significant differences among different plant life forms |
2.3 水生植物和陆生植物间SLA与LDMC的差异
本研究中的水生植物均为挺水植物,SLA和LDMC在水生和陆生植物间存在显著差异,其变异系数分别为24.4% 和21.1%。由图 3可知,水生植物SLA(148.4±20.4 cm2/g)显著小于陆生植物(210.4±56.2 cm2/g),而LDMC(0.389±0.086 g/g)则显著高于陆生植物(0.288±0.085 g/g)。

|
| 图 3 水生植物和陆生植物间SLA和LDMC的比较 Fig. 3 Comparisons of SLA and LDMC between hydrophytes and terrestrial plants Hy:水生植物Hydrophytes,Te:陆生植物Terrestrial plants小写英文字母不同者表示水生植物和陆生植物间差异显著 Different lowercase letters represent significant differences between hydrophytes and terrestrial plants |
2.4 SLA与LDMC的相关关系
综合分析19种植物SLA和LDMC的相关关系,结果显示SLA和LDMC之间呈显著负相关关系(R2=0.58),即SLA较大的物种,其LDMC较低。从不同生活型植物SLA和LDMC的散点分布(图 4)可以看出,SLA和LDMC之间在木本植物、多年生草本和1年生草本植物中均呈显著负相关关系。在3种生活型中,SLA和LDMC的相关关系在木本植物中最强,其次为多年生草本植物,在1年生草本植物中最弱。

|
| 图 4 SLA和LDMC的相关关系 Fig. 4 Correlation relationship between SLA and LDMC 黑色和灰色实线及黑色虚线分别表示木本植物、多年生草本和1年生草本SLA和LDMC的相关关系 The black and grey solid trend lines and the black dashed trend line represent the SLA-LDMC relationships of woody plants, perennial and annual herbs, respectively |
2.5 不同类型湿地间植物SLA与LDMC的差异
对于在桑沟湾湿地和红荷湿地均有分布的植物,桑沟湾湿地7种植物SLA(176.8±64.0 cm2/g)的平均值小于红荷湿地(288.3±84.8 cm2/g),而LDMC(0.352±0.104 g/g)则高于红荷湿地(0.259±0.051 g/g)。从每种植物SLA和LDMC在桑沟湾湿地和红荷湿地间的差异(图 5)可以看出,除了菖蒲外,桑沟湾湿地其余6种植物的SLA均小于红荷湿地,差异显著;桑沟湾湿地7种植物的LDMC均高于红荷湿地,差异显著。

|
|
图 5 7个物种SLA和LDMC在不同类型湿地间的比较 Fig. 5 Comparisons of SLA and LDMC of seven species in different types of wetlands Hs:葎草;Ag:艾草;Cc:小蓬草;Ac:菖蒲;Af:紫穗槐;Ic:白茅;Pa:芦苇 小写英文字母不同者表示不同类型湿地间差异显著 Hs: Humulus scandens; Ag: Artemisia argyi; Cc: Conyza canadensis; Ac: Acorus calamus; Af: Amorpha fruticosa; Ic: Imperata cylindrica; Pa: Phragmites australis Different lowercase letters represent significant differences in different types of wetlands |
为了进一步分析引起两个湿地植物间SLA和LDMC差异的环境因子,我们比较测定了桑沟湾湿地和红荷湿地土壤全氮含量的差异,结果显示桑沟湾湿地土壤全氮含量(0.150±0.018 cm2/g)显著低于红荷湿地(0.370±0.021 mg/g)。
3 讨论SLA和LDMC是植物叶片的两个主要性状,可表征植物获取资源的能力[10]。本研究中,不同物种和不同科植物间的SLA和LDMC差异显著,反映了植物对生存环境适应策略的不同。19种植物中,葎草和鸭跖草均具有较大的SLA和较低的LDMC,体现了较强的资源获取能力和较高的生产力,属于资源获取型植物,对资源丰富和湿润的生境适应能力较强;白茅和芦苇具有较小的SLA和较高的LDMC,说明其对体内养分和水分等资源的保持能力较强,比较适应资源贫瘠和干旱的生境,属于资源保守型植物[15]。在科水平上,禾本科和豆科是桑沟湾湿地分布最广的植物,两科植物具有相对较高的LDMC和较小的SLA(图 1),而这与桑沟湾湿地的气候和土壤条件密切相关。为了探讨这一问题,我们对比分析了7个物种SLA和LDMC在桑沟湾湿地和微山湖红荷湿地间的差异,结果发现桑沟湾湿地植物具有更高的LDMC和更小的SLA。桑沟湾湿地属于浅海河口湿地,其年平均降水量和年平均气温与红荷湿地差异不大,但是其年平均风速显著大于红荷湿地,且每年的大风次数较多,生长于此的植物可能受到风胁迫的影响。风胁迫不仅可以引起植物的机械损伤,而且会通过吸水的作用加速植物叶片的水分流失[22, 25]。研究发现,在风胁迫下,植物叶片表皮和角质层厚度增加,SLA显著降低,这种变化可以有效减少大风带来的机械损伤和水分散失,增强植物对大风环境的适应能力[26-27]。此外,桑沟湾湿地土壤全氮含量显著低于红荷湿地。因此,桑沟湾湿地植物较高的LDMC和较小的SLA是对该地区大风环境以及较为贫瘠土壤的一种适应策略。我们进一步将桑沟湾湿地测定数据与以往研究结果进行了对比,发现19种植物SLA数值大小处于中等偏下水平[28-29]。
诸多研究表明,SLA和LDMC在不同生活型间存在差异,相对于草本植物,木本植物具有高的LDMC和小的SLA,而多年生草本植物的LDMC要高于1年生草本植物,SLA则小于1年生草本植物[11, 29],本研究结果基本支持这一结论。在桑沟湾湿地生态系统中,草本植物处于木本植物的下方,遮阴条件下,植物为了获得更多的光照资源,叶片变薄变大,叶密度降低,导致SLA增大而LDMC降低[30-32]。本研究中,SLA和LDMC在水生植物和陆生植物间差异显著,挺水植物具有更小的SLA和更高的LDMC,这与以往的研究结果不同。以往研究认为水生植物长期处在低光和缺氧的环境中,一般具有较高的SLA和较低的LDMC[33];也有研究表明,SLA和LDMC在水生植物和陆生植物间并没有显著差异[34]。本研究中的水生植物均为挺水植物,只是根系被淹没,而叶片仍旧停留在空气中,根系周围的厌氧环境可能会阻碍植物对水分的吸收,而叶片蒸发需求保持不变,因此有可能引起生理干旱,导致植物遭受干旱胁迫,这可能是本研究中挺水植物SLA较小而LDMC较高的原因[35]。
植物对环境的适应往往表现为一系列叶性状的组合[36]。本研究也发现,SLA和LDMC之间在不同生活型中均呈显著负相关关系,这一关系在植物群落中普遍存在[11-12],二者可一起作为植物资源利用分类轴上划分植物种类的最佳变量来筛选用于恢复特定湿地功能的植物,进而有效地对湿地资源进行保护和合理开发。
4 结论本研究结果表明,桑沟湾湿地不同科植物间以及不同生活型植物间SLA和LDMC差异显著,说明该地区植物可通过对生存环境采取不同的适应策略以实现对环境资源利用效率的最大化和植物生长的最大化。而相对于微山湖红荷湿地来说,桑沟湾湿地植物较高的LDMC和较小的SLA是对该地区大风环境以及较为贫瘠土壤的一种适应策略。
| [1] |
吕金霞, 蒋卫国, 王文杰, 陈坤, 邓越, 陈征, 荔琢. 近30年来京津冀地区湿地景观变化及其驱动因素[J]. 生态学报, 2018, 38(12): 4492-4503. DOI:10.5846/stxb201712232309 LYU J X, JIANG W G, WANG W J, CHEN K, DENG Y, CHEN Z, LI Z. Wetland landscape pattern change and its driving forces in Beijing-Tianjin-Hebei region in recent 30 years[J]. Acta Ecologica Sinica, 2018, 38(12): 4492-4503. DOI:10.5846/stxb201712232309 |
| [2] |
雷昆, 张明祥. 中国的湿地资源及其保护建议[J]. 湿地科学, 2005, 3(2): 81-86. DOI:10.3969/j.issn.1672-5948.2005.02.001 LEI K, ZHANG M X. The wetland resources in China and the conservation advices[J]. Wetland Science, 2005, 3(2): 81-86. DOI:10.3969/j.issn.1672-5948.2005.02.001 |
| [3] |
张慧, 李智, 刘光, 祝炜平. 中国城市湿地研究进展[J]. 湿地科学, 2016, 14(1): 103-107. DOI:10.13248/j.cnki.wetlandsci.2016.01.016 ZHANG H, LI Z, LIU G, ZHU W P. Advances in the study of urban wetlands in China[J]. Wetland Science, 2016, 14(1): 103-107. DOI:10.13248/j.cnki.wetlandsci.2016.01.016 |
| [4] |
MOOR H, RYDIN H, HYLANDER K, NILSSON M B, LINDBORG R, NORBERG J. Towards a trait-based ecology of wetland vegetation[J]. Journal of Ecology, 2017, 105: 1623-1635. DOI:10.1111/1365-2745.12734 |
| [5] |
HU Y K, LIU G F, PAN X, SONG Y B, DONG M, CORNELISSEN J H C. Abundance-weighted plant functional trait variation differs between terrestrial and wetland habitats along wide climatic gradients[J]. Science China Life Sciences, 2020, 63: 1-13. DOI:10.1007/s11427-020-1766-1 |
| [6] |
PAN Y J, CIERAAD E, ARMSTRONG J, ARMSTRONG W, CLARKSON B R, COLMER T D, PEDERSEN O, VISSER E J W, VOESENEK L A C J, BODEGOM P M V. Global patterns of the leaf economics spectrum in wetlands[J]. Nature Communications, 2020, 11: 1-9. DOI:10.1038/s41467-020-18354-3 |
| [7] |
CARVALHO F, BROWN K A, WALLER M P, BOOM A. Leaf traits interact with management and water table to modulate ecosystem properties in fen peatlands[J]. Plant and Soil, 2019, 441: 331-347. DOI:10.1007/s11104-019-04126-6 |
| [8] |
KUNERT N, ZAILAA J. Specific leaf area for five tropical tree species growing in different tree species mixtures in Central Panama[J]. New Forests, 2019, 50: 873-890. DOI:10.1007/s11056-019-09706-z |
| [9] |
WRIGHT I J, REICH P B, WESTOBY M, ACKERLY D D, BARUCH Z, BONGERS F, CAVENDER-BARES J, CHAPIN T, CORNELISSEN J H C, DIEMER M, FELXAS J, GARNIER E, GROOM H C, GULIAS J, KIKOSAKA K, LAMONT B B, LEE T, LEE W, LUSK C, MIDGLEY J J, NAVAS M, NⅡNEMETS ü, OLEKSYN J, OSADA N, POORTER H, PRIOR P L, PYANKOV V I, ROUMET C, THOMAS S C, TJOELKER M G, VENEKLASS E J, VILLAR R. The worldwide leaf economics spectrum[J]. Nature, 2004, 428: 821-827. DOI:10.1038/nature02403 |
| [10] |
WILSON P J, THOMPSON K, HODGSON J G. Specific leaf area and leaf dry matter content as alternative predictors of plant strategies[J]. New Phytologist, 1999, 143: 155-162. DOI:10.1046/j.1469-8137.1999.00427.x |
| [11] |
李玉霖, 崔建垣, 苏永中. 不同沙丘生境主要植物比叶面积和叶干物质含量的比较[J]. 生态学报, 2005, 25(2): 304-311. DOI:10.3321/j.issn:1000-0933.2005.02.019 LI Y L, CUI J H, SU Y Z. Specific leaf area and leaf dry matter content of some plants in different dune habitats[J]. Acta Ecologica Sinica, 2005, 25(2): 304-311. DOI:10.3321/j.issn:1000-0933.2005.02.019 |
| [12] |
黄雅茹, 马迎宾, 苏智, 李炜, 张格, 赵英铭, 董雪, 刘明虎. 乌兰布和沙漠绿洲北抗杨家系6个无性系叶片性状研究[J]. 西北林学院学报, 2019, 34(3): 86-90. DOI:10.3969/j.issn.1001-7461.2019.03.13.j.issn.1001-7461.2019.03.13 HUANG Y R, MA Y B, SU Z, LI W, ZHANG G, ZHAO Y M, DONG X, LIU M H. Leaf traits of six clones of Populus deltoides cl. Beikang family in Ulan Buh Desert Oasis[J]. Journal of Northwest Forestry University, 2019, 34(3): 86-90. DOI:10.3969/j.issn.1001-7461.2019.03.13.j.issn.1001-7461.2019.03.13 |
| [13] |
HULSHOF C M, SWENSON N G. Variation in leaf functional trait values within and across individuals and species: an example from a Costa Rican dry forest[J]. Functional Ecology, 2010, 24: 217-223. DOI:10.1111/j.1365-2435.2009.01614.x |
| [14] |
张剑, 包雅兰, 宿力, 王利平, 陆静雯, 曹建军. 敦煌阳关湿地芦苇叶性状对土壤水分的响应[J]. 生态学报, 2019, 39(20): 7670-7678. DOI:10.5846/stxb201808231976 ZHANG J, BAO Y L, SU L, WANG L P, LU J W, CAO J J. Response of Phragmites australis leaf traits to soil moisture in Yangguan wetland, Dunhuang[J]. Acta Ecologica Sinica, 2019, 39(20): 7670-7678. DOI:10.5846/stxb201808231976 |
| [15] |
POORTER L, BONGERS F. Leaf traits are good predictors of plant performance across 53 rain forest species[J]. Ecology, 2006, 87: 1733-1743. DOI:10.1890/0012-9658(2006)87[1733:LTAGPO]2.0.CO;2 |
| [16] |
钟巧连, 刘立斌, 许鑫, 杨勇, 郭银明, 许海洋, 蔡先立, 倪健. 黔中喀斯特木本植物功能性状变异及其适应策略[J]. 植物生态学报, 2018, 42(5): 562-572. DOI:10.17521/cjpe.2017.0270 ZHONG Q L, LIU L B, XU X, YANG Y, GUO Y M, XU H Y, CAI X L, NI J. Variations of plant functional traits and adaptive strategy of woody species in a karst forest of central Guizhou Province, southwestern China[J]. Chinese Journal of Plant Ecology, 2018, 42(5): 562-572. DOI:10.17521/cjpe.2017.0270 |
| [17] |
杨建军, 苏文华, 王玲玲, 杨锐, 杨波, 张博睿. 高山栲叶性状种内变异及其与环境因子的关系[J]. 广东农业科学, 2015, 42(12): 152-158. DOI:10.16768/j.issn.1004-874X.2015.12.030 YANG J J, SU W H, WANG L L, YANG R, YANG B, ZHANG B R. Intraspecific variations of Castanopsis delavayi leaf traits and their relationship with environmental factors[J]. Guangdong Agricultural Sciecnes, 2015, 42(12): 152-158. DOI:10.16768/j.issn.1004-874X.2015.12.030 |
| [18] |
李颖, 姚婧, 杨松, 侯继华. 东灵山辽东栎林主要树种叶功能性状差异性研究[J]. 广东农业科学, 2012, 39(23): 159-162, 171. DOI:10.3969/j.issn.1004-874X.2012.23.050 LI Y, YAO J, YANG S, HOU J H. Trait differences research on leaf function of liaoning oak forest main species in Dongling mountain[J]. Guangdong Agricultural Sciences, 2012, 39(23): 159-162, 171. DOI:10.3969/j.issn.1004-874X.2012.23.050 |
| [19] |
魏丽萍, 侯继华, 蒋思思. 阔叶红松林两个主要树种叶功能性状随树高变化[J]. 广东农业科学, 2014, 41(12): 55-58. DOI:10.16768/j.issn.1004-874X.2014.12.013 WEI L P, HOU J H, JIANG S S. Changes of leaf functional traits of two main species along tree height in broad-leaved Korean pine forest[J]. Guangdong Agricultural Sciences, 2014, 41(12): 55-58. DOI:10.16768/j.issn.1004-874X.2014.12.013 |
| [20] |
王茜, 肖辉明, 刘彩玲, 张宗国. 荣成市国家城市湿地公园现状及保护措施[J]. 山东林业科技, 2006(3): 48-49. DOI:10.3969/j.issn.1002-2724.2006.03.022 WANG Q, XIAO H M, LIU C L, ZHANG Z G. Current situation and conservation measures of Rongcheng National Urban Wetland Park[J]. Journal of Shandong Forestry Science and Technology, 2006(3): 48-49. DOI:10.3969/j.issn.1002-2724.2006.03.022 |
| [21] |
SMITH V C, ENNOS A R. The effects of air flow and stem flexure on the mechanical and hydraulic properties of the stems of sunflowers Helianthus annuus L[J]. Journal of Experimental Botany, 2003, 54(383): 845-849. DOI:10.1093/jxb/erg068 |
| [22] |
张鹏. 海岸梯度上黑松针叶性状的适应性研究[D]. 北京: 中国林业科学研究院, 2018. ZHANG P. Study on adaptability in needle traits of Pinus thunbergii along coastal-inland gradient[D]. Beijing: Chinese Academy of Forestry, 2018. |
| [23] |
WU T G, ZHANG P, ZHANG L, WANG G G, YU M. Morphological response of eight Quercus species to simulated wind load[J]. PLOS ONE, 2016, 11(9): 1-14. DOI:10.1371/journal.Pone.0163613 |
| [24] |
KJELDAHL J. A new method for the determination of nitrogen in organic matter[J]. Zeitschrift fur Analytische Chemie Fresenius, 1883, 22: 366-382. DOI:10.1007/BF01338151 |
| [25] |
ANTEN N P R, ALCALA-HERRERA R, SCHIEVING F, YUSUKE O. Wind and mechanical stimuli differentially affect leaf traits in Plantago major[J]. New Phytologist, 2010, 188(2): 554-564. DOI:10.1111/j.1469-8137.2010.03379.x |
| [26] |
VOGEL S. Leaves in the lowest and highest wind: temperature, force and shape[J]. New Phytologist, 2009, 183(1): 13-26. DOI:10.1111/j.1469-8137.2009.02854.x |
| [27] |
古力米热热孜, 赵晓英, 原慧. 模拟长期大风胁迫对霸王叶解剖结构特征的影响[J]. 西北植物学报, 2012, 32(10): 2047-2052. DOI:10.3969/j.issn.1000-4025.2012.10.018 GULIMIRE R, ZHAO X Y, YUAN H. Leaf anatomical structure of Zygophyllum xanthoxylum under chronic high wind[J]. Acta Botanica Boreali-Occidentalia Sinica, 2012, 32(10): 2047-2052. DOI:10.3969/j.issn.1000-4025.2012.10.018 |
| [28] |
刘贤娴, 李俊清. 北京野鸭湖湿地植物叶功能性状研究[J]. 安徽农业科学, 2008, 36(20): 8406-8409. DOI:10.13989/j.cnki.0517-6611.2008.20.019 LIU X X, LI J Q. Study on the functional traits of plant leaf in Beijing Yeyahu wetland[J]. Journal of Anhui Agricultural Sciences, 2008, 36(20): 8406-8409. DOI:10.13989/j.cnki.0517-6611.2008.20.019 |
| [29] |
POORTER H, NⅡNEMETS ü, POORTER L, WRIGHT I J, VILLAR R. Causes and consequences of variation in leaf mass per area(LMA): a meta-analysis[J]. New Phytologist, 2009, 182: 565-588. DOI:10.1111/j.1469-8137.2009.02830.x |
| [30] |
LUSK C H, REICH P B, MONTGOMERY R A, ACKERLY D D, CAVENDER-BARES J. Why are evergreen leaves so contrary about shade?[J]. Trends in Ecology and Evolution, 2008, 23(6): 299-303. DOI:10.1016/j.tree.2008.02.006 |
| [31] |
毛伟, 李玉霖, 赵学勇, 黄迎新, 王少昆. 科尔沁沙地灌丛内外草本植物狗尾草叶性状的比较研究[J]. 草业学报, 2009, 18(6): 144-150. DOI:10.11686/cyxb20090619 MAO W, LI Y L, ZHAO X Y, HUANG Y X, WANG S K. Effect of Caragana microphylla on leaf traits of Setarria viridis in Horqin sandy land[J]. Acta Prataculturae Sinica, 2009, 18(6): 144-150. DOI:10.11686/cyxb20090619 |
| [32] |
刘明秀, 梁国鲁. 植物比叶质量研究进展[J]. 植物生态学报, 2016, 40(8): 847-860. DOI:10.17521/cjpe.2015.0428 LIU M X, LIANG G L. Research progress on leaf mass per area[J]. Chinese Journal of Plant Ecology, 2016, 40(8): 847-860. DOI:10.17521/cjpe.2015.0428 |
| [33] |
PIERCE S, BRUSA G, SARTORI M, CERABOLINI B E L. Combined use of leaf size and economics traits allows direct comparison of hydrophyte and terrestrial herbaceous adaptive strategies[J]. Annals of Botany, 2012, 109: 1047-1053. DOI:10.1093/aob/mcs021 |
| [34] |
张丽霞. 基于功能性状的水生植物及邻近陆生植物的生态策略[D]. 西安: 西北大学, 2017. ZHANG L X. Economics trade-offs of hydrophytes and adjacent terrestrial herbaceous plants based on plant functional traits[D]. Xi'an: Northwest University, 2017. |
| [35] |
PAN Y J, CIERAAD E, BODEGOM P M. Are ecophysiological adaptive traits decoupled from leaf economics traits in wetlands?[J]. Functional Ecology, 2019, 33(7): 1202-1210. DOI:10.1111/1365-2435.13329 |
| [36] |
GRIME J P. Trait convergence and trait divergence in herbaceous plant communities: Mechanisms and consequences[J]. Journal of Vegetation Science, 2006, 17(2): 255-260. DOI:10.1111/j.1654-1103.2006.tb02444.x |