 2021, Vol. 48
2021, Vol. 48文章信息
引用本文 |

基金项目
- 广东省现代农业产业技术体系茶叶创新团队项目(2020KJ120);中国科学院科技服务网络计划区域重点项目(KFJ-STS-QYZX-093)
作者简介
- 傅秀敏(1986—),女,博士,副研究员,研究方向为园艺植物品质形成与调控,E-mail:fuxiumin@scbg.ac.cn.
文章历史
- 收稿日期:2021-03-03

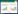

2. 广东省农业科学院茶叶研究所,广东 广州 510640
2. Tea Research Institute, Guangdong Academy of Agricultural Sciences, Guangzhou 510640, China
植物类胡萝卜素主要分布于植物细胞的叶绿体和有色体两个亚细胞器中。在叶绿体中,类胡萝卜素主要发挥光捕获作用和光破坏防御功能[1-2],其组成及比例比较保守[3-4],主要积累叶黄质(Lutein)、β- 胡萝卜素(β-carotene)、堇菜黄素(Violaxanthin)、花药黄质(Antheraxanthin)、玉米黄质(Zeaxanthin)和新黄质(Neoxanthin)。在有色体中,类胡萝卜素使细胞和相应组织呈红色、橙色及黄色,主要起到吸引昆虫进行授粉或传播种子的作用[5]。相比叶绿体,有色体中类胡萝卜素的种类较为丰富,且各组分间的比例与物种相关[6-10]。
目前研究类胡萝卜素的合成及代谢主要集中在各物种的果实及花中,包括采前和采后阶段[11]。在绿色组织方面的研究主要集中于模式植物拟南芥,主要关注其生长阶段类胡萝卜素在光合作用、生长发育及激素调控方面的作用,并利用其突变体对类胡萝卜素途径中的关键基因进行体内的功能验证[4]。茶树为我国重要园艺经济作物之一,茶叶品质是茶叶产品的核心价值,许多特征性代谢物包括多酚、氨基酸(尤其是茶氨酸)、香气和色素类物质(主要为叶绿素和类胡萝卜素)对其品质具有重要贡献价值。类胡萝卜素物质在茶叶中除贡献茶叶叶底颜色外,还是茶叶重要香气物质的前体物,尤其在茶叶加工过程中类胡萝卜素会经酶促或非酶促反应降解生成具有花香味的香气物质。已有研究表明,类胡萝卜素的含量和组成及其降解产物决定茶叶品质[12-13],其合成代谢与调控研究备受关注。
茶叶类胡萝卜素在生长发育阶段的形成机制与拟南芥类似,但也存在特殊性,如茶叶是多年生常绿木本植物,具有不同采收季节。此外,人们在食用茶叶前,需经过一系列加工过程,且不同加工工艺可制成不同茶类。在模式植物拟南芥中,目前尚无采后加工对类胡萝卜素合成代谢影响的研究,采后绿色组织中类胡萝卜素的合成及代谢调控机制还有待进一步阐明。茶叶实际生产过程贯穿茶树生长过程(采前阶段)和茶叶加工过程(采后阶段),茶叶类胡萝卜素的研究将为深入了解采后绿色组织中类胡萝卜素的合成及代谢调控机制提供基础。鉴于类胡萝卜素对茶叶品质的重要贡献,本文综述了茶树资源(绿色正常种和黄化或白化突变种)、茶树生长环境(光和温度)、农艺措施(遮阴)及采后加工工艺(萎凋和干燥)对茶叶类胡萝卜素合成及代谢的影响规律与调控机制,最后探讨茶叶类胡萝卜素的未来研究方向,为今后茶叶品质改良提供一定的理论依据。
1 茶叶类胡萝卜素的生物合成调控 1.1 胡萝卜素在不同茶树种质资源中的积累机制差异茶树种质资源丰富,除绿色品种外,还存在一些黄化或白化的突变品种。这些黄化或白化突变品种的叶片具有高氨基酸、低多酚的特性,其制成的茶叶品质较高,被广泛栽培[14-15]。由于黄化或白化茶树品种的叶片不能积累叶绿素,类胡萝卜素主要贡献了这些突变品种的色泽,茶叶中许多类胡萝卜素研究都以这些突变品种作为研究材料。Wang等[16]比较了正常绿色品种龙井43和黄化品种中黄2号的类胡萝卜素含量,发现黄化品种中β- 胡萝卜素、叶黄质含量显著低于绿色品种,仅为绿色品种中含量的88.6%、48.8%[16],通过进一步电镜观察发现黄化种的叶绿体基粒结构已发生降解。Feng等[17]比较了正常绿色品种福鼎大白茶和4个黄化品种安吉白茶、黄金芽、天台黄茶和郁金香的类胡萝卜素含量,发现4个黄化种的总类胡萝卜素含量都显著低于绿色正常种。除玉米黄质在4个黄化品种中含量高于绿色对照种外,叶黄质、β- 胡萝卜素和堇菜黄素含量均显著低于绿色品种。李明等[18]同样发现光照敏感型黄化茶叶御金香和温度敏感型白化茶叶白叶一号的类胡萝卜素总含量均显著低于绿色对照品种福鼎大白茶。可见,黄化或白化突变种伴随着叶绿体结构的瓦解,不但叶绿素含量下降,类胡萝卜素含量也显著下降。
在植物类胡萝卜素研究中,八氢番茄红素脱氢酶PDS功能的缺失或基因的低表达水平导致植株黄化白化[19]。李娜娜等[20]比较了茶树CsPDS在黄化品种黄金芽和正常绿色品种福鼎大白茶中的转录水平,发现其表达水平在黄金芽中并未受到抑制,推测该基因不是黄金芽黄化和类胡萝卜素含量低的主要原因。Li等[21]进一步比较了黄金芽和福鼎大白茶中茶叶类胡萝卜素合成途径上游1- 脱氧-D- 木酮糖-5- 磷酸合酶基因DXS 3个成员(DXS1、DXS2和DXS3)和下游合成基因包括八氢番茄红素合成酶(PSY)、八氢番茄红素去饱和酶(PDS)、ζ- 胡萝卜素去饱和酶(ZDS)、番茄红素ε环化酶(LCYE)、番茄红素β环化酶(LCYB)、胡萝卜素ε羟化酶(LUT1)、玉米黄质环氧酶(ZEP)和堇菜黄素脱环氧酶(VDE)的转录水平,发现只有DXS的3个成员在黄化种中的表达量极显著低于绿色正常种,推测其低表达量与黄化品种中类胡萝卜素含量低有关。Zheng等[22]以茶树品种水仙及其黄化突变体为试材,也发现黄化突变种的类胡萝卜素含量显著低于绿色品种对照,并通过转录组学手段发现类胡萝卜素合成上游基因CsDXS基因和香叶基香叶基二磷酸合酶CsGGPS基因的表达量显著低于绿色品种对照,其表达量水平也与类胡萝卜素含量呈正相关。综上分析可知,尽管类胡萝卜素是黄化突变种中的主要色泽贡献物质,但其在黄化种中含量低于绿色种。此外,类胡萝卜素合成的上游基因CsDXS和CsGGPS基因的表达量低可能是黄化品种中类胡萝卜素积累少的部分原因(图 1)。

|
| 图 1 茶树采前及采后类胡萝卜素的合成及代谢调控 Fig. 1 The biosynthesis and metabolism regulation of carotenoids in tea plants during the stage of growth and post-harvest processing BCH: 胡萝卜素β羟化酶;CMK: 4-(胞苷5’-二磷酸)-2-C-甲基-D-赤藓糖醇激酶;CCD: 类胡萝卜素裂解双加氧酶;CRTISO: 胡萝卜素异构酶;DXS: 1- 脱氧-D- 木酮糖-5- 磷酸合成酶;DXR: 1- 脱氧-D- 木酮糖-5- 磷酸还原异构酶;ECH: 胡萝卜素ε羟化酶;GGPS: 香叶基香叶基二磷酸合酶;HDR: 4- 羟基-3- 甲基丁-2- 烯-1- 基二磷酸还原酶;LCYB: 番茄红素β环化酶;LCYE: 番茄红素ε环化酶;MCT: 2-C- 甲基-D- 赤藓糖醇4- 磷酸胞苷转移酶;NCED: 9- 顺- 环氧类胡萝卜素双加氧酶;NXS: 新黄质合成酶;PSY: 八氢番茄红素合成酶;PDS: 八氢番茄红素去饱和酶;VDE: 堇菜黄素脱环氧酶;ZDS: ζ- 胡萝卜素去饱和酶;ZEP: 玉米黄质环氧酶;ZISO: ζ- 胡萝卜素异构酶 BCH: β-carotene hydroxylase; CMK: 4-(cytidine 5'-diphospho)-2-Cmethyl-D-erythritol kinase activity; CCD: carotenoid cleavage oxygenases; CRTISO: carotene isomerase; DXS: 1-deoxy-D-xylulose 5-phosphate synthase; DXR: 1-deoxy-D-xylulose 5-phosphate reductoisomerase; ECH: ε-carotene hydroxylase; GGPS: geranylgeranyl diphosphate synthase; HDR: 4-hydroxy-3-methylbut-2-en-1-yl diphosphate reductase; LCYB: lycopene β-cyclase; LCYE: lycopene ε-cyclase; MCT: 2-C-methylD-erythritol 4-phosphate cytidylyltransferase; NCED: 9-cis-epoxycarotenoid dioxygenase; NXS: neoxanthin synthase; PSY: phytoene synthase; PDS: phytoene desaturase; VDE: violaxanthin de-epoxidase; ZDS: ζ-carotene desaturase; ZEP: zeaxanthin epoxidase; ZISO: ζ-carotene isomerase. |
虽然类胡萝卜素的组成在绿色组织中比较保守,但其含量在不同物种或品种中不同[23-24]。Wang等[25]比较了来自4个国家(中国、日本、肯尼亚和格鲁吉亚)的119份茶树种质资源(包括79份Camellia. sinensis var. sinensis、18份C. sinensis var. assamica和22份C. sinensis var. pubilimba)的类胡萝卜素含量,发现不同种质资源间类胡萝卜素含量分布存在显著差异,β- 胡萝卜素和叶黄素含量最高的种质资源分别为含量最低种质资源的42.1倍和16.3倍。Wei等[26]分析了我国3个产区31个茶树品种的类胡萝卜素含量,这些品种类胡萝卜素总量为324.8~528.8 μg/g,且遗传因素对类胡萝卜素含量的影响比栽培环境更大。陈丽等[27]发现37个不同适制性茶树品种的类胡萝卜素组成及含量存在显著差异,而同一适制性茶树品种间具有相同的分布规律。上述研究表明,类胡萝卜素在茶叶上的分布具有丰富的资源多样性,且在不同适制性的茶叶品种上具有一定关联性。
茶树主要分为中国种和阿萨姆种两大类。Borthakur等[28]研究发现中国小叶种(龙井43)茶叶组织中类胡萝卜素总含量高于阿萨姆大叶种(玉兰)。Venkatakrishna[29]发现类胡萝素含量在中国种(Thea sinensis)中的含量是阿萨姆种(Thea assamica)两倍左右。Ravichandran等[30]也发现中国种茶叶的类胡萝卜素含量高于阿萨姆种,表明如果增加茶叶内源性类胡萝卜素的含量会提升茶叶香气风味品质指数,且通过合理施肥会促进类胡萝卜素积累。综上可知,中国种茶叶的类胡萝卜素含量普遍高于阿萨姆种,其类胡萝卜素含量差异积累的机制尚不明确。
除种质资源影响外,不同成熟阶段的茶树叶片中类胡萝卜素含量存在差异,这点与其他植物类似[31]。Venkatakrishna等[32]研究发现类胡萝卜素含量和组成在茶树叶片不同发育阶段不同;类胡萝卜素含量随叶片成熟度增加呈增加趋势,第一叶的类胡萝卜素含量每100 g仅为25.42 mg,而成熟叶中类胡萝卜素含量每100 g达到126.08 mg。除含量差异外,类胡萝卜素组成也存在差异;叶黄素比例在幼嫩叶片中较高,从幼嫩叶组织的胡萝卜素和叶黄素比值(1∶2.5)降到成熟叶片的1∶1.3。不同成熟度的茶叶加工后呈现不同香型,类胡萝卜素来源的香气物质可能是其香型的重要影响因子。而目前成熟度对茶叶类胡萝卜素含量及组成的影响机制还未明确,有待进一步研究。
1.2 茶叶采前阶段外界环境因子对类胡萝卜素积累的影响1.2.1 光 已有大量研究证实,植物类胡萝卜素的积累可受光调控[11]。在茶树中,正常的绿色茶树品种在进行遮阴处理后,叶片颜色变得更绿,类胡萝卜素含量也会积累更多。Sonobe等[33]采用不同遮阴处理(0%、35%、75% 和90%)发现,类胡萝卜素含量随遮阴程度加重而增加。此外,研究发现更多的类胡萝卜素积累会对加工后茶的风味产生影响。例如,日本的Kabuse-cha茶具有一种特征性香气物质‘Ooika’,Kawakam等[34]研究发现,‘Ooika’可能是一种类胡萝卜素来源的香气物质,这是由于Kabuse-cha茶原料来自遮阴处理的鲜叶加工而成,该茶含有较高的类胡萝卜素含量及类胡萝卜素来源的香气物质。除光照强度影响茶叶的类胡萝卜素积累外,光质也会参与影响茶叶类胡萝卜素积累,如单一蓝光处理会促进茶叶类胡萝卜素含量增加[35]。
光是影响光敏感型茶树叶片表型及代谢物积累的重要环境因子,光照对类胡萝卜素积累调控机制的研究主要以光敏感型茶树为材料。李明等[18]研究发现,茶树光敏感型品系(黄-2、黄-8和黄-13)的类胡萝卜素含量显著低于绿色对照种(福鼎大白茶);茶树光敏感型品系在进行遮阴处理后可显著提高类胡萝卜素含量,但仍低于正常绿色种;遮阴处理显著提高茶树光敏感型品系的β- 胡萝卜素和新黄质含量,对新黄质含量无显著影响,但显著降低紫黄质(堇菜黄素)含量。范延艮[36]研究发现,光敏感型黄化品种黄金芽在遮阴处理后堇菜黄素含量上升,而环氧玉米黄质含量降低,表明遮阴后叶黄素循环降低;对黄金芽进行遮阴处理后,同样发现遮阴后黄金芽叶片返绿,类胡萝卜素含量增加;进一步通过转录组和基因表达水平分析,发现类胡萝卜素合成和代谢相关基因PSY、PDS、ZDS、LCYE、LCYB、ZEP、VDE以及胡萝卜素ε羟化酶(BCH)、胡萝卜素ε羟化酶(ECH)和类胡萝卜素裂解双加氧酶(CCD1、CCD4和CCD8)的转录水平在遮阴后均被上调,与类胡萝卜素含量呈正相关[37]。
1.2.2 温度 温度是影响温度敏感型茶树幼叶表型及代谢物积累的重要环境因子之一,研究温度对茶叶类胡萝卜素积累的影响主要以温度敏感型茶树品种为材料[38]。小雪芽为温度敏感型茶树品种,在生长温度15℃条件下新抽出的嫩梢为白色,是叶绿素和类胡萝卜素缺乏导致[39]。Du等[40]研究发现,小雪芽在15℃生长环境中叶片的类胡萝卜素含量极显著低于25℃下(仅为其3%);而在绿色对照种中,虽然低温会抑制其类胡萝卜素的积累,但在15℃下类胡萝卜素含量的下降幅度为25℃下的50% 左右。通过叶绿素和类胡萝卜素相关基因的表达分析,发现参与叶黄素循环的VDE基因在15℃低温处理的小雪芽品种中,其表达量被显著抑制。邓婷婷[41]比较了另一个温度敏感型茶树安吉白茶在15、19、23℃处理条件下类胡萝卜素相关基因的表达量,发现PSY、PDS3、ZDS、LCYB、LCYE和VDE的表达量在15℃条件下的白化阶段均受到抑制。上述研究表明,温度是影响茶叶类胡萝卜素积累的一个关键因子(图 1),低温会通过抑制类胡萝卜素合成关键基因的表达量促使类胡萝卜素含量下降,尤其是对温度敏感型的茶树。
1.2.3 季节 根据采收季节的不同,茶叶可分为春茶、夏茶、秋茶及冬茶,一般以春茶的品质最佳。Borthakur等[28]发现浙江两个茶树品种玉兰和龙井43叶片在不同季节中类胡萝卜素含量不同,且两个品种的类胡萝卜素含量随季节变化呈不同的变化规律;玉兰在春季叶片的类胡萝卜素含量最高、夏季茶叶含量最低,而龙井43在夏季茶叶中的类胡萝卜素含量最高、在春季叶片含量最低;进一步对PSY基因的表达量分析发现,该基因的转录水平在两个品种中均随不同季节变化,且与类胡萝卜素含量呈现较好的相关性(图 1)。Hazarika等对印度4个具有遗传背景差异的茶树品种〔C. sinensis(L.)O. Kuntze;C. assamica(Masters)Weitht;C. assamica ‘Cambod’和C. sinensis, hybrid〕进行不同季节间的类胡萝卜素含量比较研究,也发现类胡萝卜素含量随季节变化而变化,且均在春季茶叶(4月)中含量最高[42]。此研究结果表明,该4个品种均具有相同的季节响应模式,这与Borthakur等[28]研究发现的季节响应模式略有不同。这可能与两个研究选取茶树样本种植区域的环境或品种差异相关。在环境方面,印度和我国浙江两个地方纬度相差较大,四季气候环境存在显著差异。印度为热带季风气候,4月平均温度达22~35℃,而浙江为亚热带季风气候,春季茶(4月)平均温度为14~24℃,两地约相差10℃。光照和温度是不同季节间的主要差异因素,两者如何协同调控茶叶类胡萝卜素积累的分子机制有待进一步研究。
2 茶叶类胡萝卜素的代谢调控植物体内类胡萝卜素可被氧化裂解生成脱辅基类胡萝卜素,如9- 顺- 环氧类胡萝卜素(9-顺- 堇菜黄素和9- 顺- 新黄质)经9- 顺- 环氧类胡萝卜素双加氧酶NCEDs裂解等步骤产生脱落酸ABA;β- 胡萝卜素经CCD7和CCD8氧化裂解产生独脚金内酯;番茄红素、β- 胡萝卜素和α- 胡萝卜素又可被CCD1和CCD4氧化裂解产生各种香气物质(如假紫罗酮、β- 紫罗酮和α- 紫罗酮等)[43]。由于类胡萝卜素降解的香气产物如β- 紫罗酮和大马士酮(Damascenone)的气味感知阈值极低,分别为0.2、0.004 μg/L,对茶叶品质具有重要贡献[44]。目前茶叶中类胡萝卜素代谢路径的研究主要集中在类胡萝卜素氧化裂解生成香气物质通路,主要包括加工过程中的酶促反应和非酶促反应[45]。
2.1 茶叶采后加工过程中类胡萝卜素的酶促反应降解目前很多研究已明确指出,在制茶过程中尤其是萎凋和发酵阶段,类胡萝卜素是形成茶叶香气的重要前提物质[46]。研究发现,堇菜黄素和新黄质含量在茶叶萎凋过程中急剧下降,而β-胡萝卜素和叶黄质含量则在发酵过程显著减少,并提出红茶香气的形成与类胡萝卜素降解密切相关。类胡萝卜素降解主要发生在红茶发酵过程中,降解生成系列香气物质,如贡献红茶香气的重要物质β- 紫罗酮[42, 45]。Sanderson等[47]通过模拟法实验证实,与氧化的儿茶素物质一起进行干燥时,β- 胡萝卜素会降解生成β- 紫罗酮。各种氧化酶和裂解酶会参与茶叶发酵阶段中类胡萝卜素的降解。有研究者从日本的秋季茶叶中分离获得类胡萝卜素裂解酶CCD,该酶最佳酶活温度接近70℃,最佳pH值为7.4;并发现春季茶叶中的CCD活性与秋季茶叶中的酶活存在差异[48]。Felfe等[49]发现,CCD蛋白在茶叶体内存在二级磷酸化修饰,秋季茶CCD蛋白的磷酸化水平高于春季茶,可能导致春季茶分离到的CCD蛋白(Km=9.45 μmol/L)和秋季茶的CCD蛋白(Km=17.16 μmol/L)呈现不同的酶活特性。今后研究可进一步探讨CCD蛋白的磷酸化修饰不同是否会引起春茶和秋茶在茶叶加工过程中β- 紫罗酮的积累差异。
植物中的CCD有许多基因家族成员,它们具有不同功能。体内及体外功能验证表明,在大部分植物中,CCD1、CCD4和CCD7具有氧化裂解多种类胡萝卜素生成具有花果香的β- 紫罗酮、α- 紫罗酮、香叶基丙酮和二氢海葵内酯等香气物质[43, 50-52]的功能。王赞等[53]通过同源克隆及转录水平分析,发现茶叶中的CCD1和CCD4基因表达水平具有组织特异性,并在乌龙茶做青过程中其转录水平发生变化,由此推测这两个基因可能参与了乌龙茶花果香味的形成。Wang等[54]监测了茶叶萎凋过程中β- 紫罗酮的含量,根据其含量与CCD基因表达的相关性筛选到CCD4成员;在β- 胡萝卜素积累的菌株中表达该基因,发现该菌株可以降解β- 胡萝卜素形成β- 紫罗酮,推测该成员具有以β- 胡萝卜素为底物催化生成β- 紫罗酮的功能;进一步研究发现,该基因的转录水平受温度和机械伤胁迫调控,但不受失水胁迫诱导。该研究团队随后又筛选到可受失水胁迫响应的CsCCD1a成员,同样具有催化β-胡萝卜素形成β- 紫罗酮的作用,推测CCD1a与CCD4基因成员在茶叶萎凋过程中共同参与裂解β- 胡萝卜素生成β- 紫罗酮,最终贡献了茶叶中的花果香味[55]。上述研究结果表明,茶叶中的CCD参与采后茶叶类胡萝卜素裂解生成香气的作用(图 1)。除CCD1和CCD4外,CCD7也被报道具有裂解β- 胡萝卜素生成β- 紫罗酮的作用[43],然而茶叶中CCD7的功能尚不清楚,有待于进一步研究。
2.2 茶叶采后加工过程中类胡萝卜素的非酶促反应降解除酶促反应外,高温条件下类胡萝卜素会发生非酶促反应进而降解。干燥是制茶工艺中的重要工序。干燥过程温度高,茶叶含水量急剧降至约3%,酶促反应也被终止,同时类胡萝卜素(尤其是胡萝卜素和叶黄质)含量也显著降低[56]。此外,研究表明茶叶在高温冲泡过程中会损失约10% 的类胡萝卜素[29]。日本学者对β- 胡萝卜素单独进行加热后发现其可形成10种香气物质,包括二氢猕猴桃内酯和β- 紫罗酮[57]。Kanasawud等[58]发现β- 胡萝卜素在97 ℃溶液中放置3 h后可降解生成50多种香气物质。上述研究表明,在茶叶加工过程的干燥工序和高温冲泡茶叶过程中,各种类胡萝卜素会被裂解成β-紫罗酮、α- 紫罗酮、芳樟醇、二氢猕猴桃内酯和茶香螺酮等萜烯类香气物质。这些香气物质贡献了部分茶叶香气中的果香味,如β- 紫罗酮和二氢猕猴桃内酯是乌龙茶和红茶花果香的重要贡献因子[59]。
大马士酮是类胡萝卜素降解形成的另一重要香气物质,常见于加工食品和饮料中,为酒精饮料中的风味成分,常见于葡萄酒中[60]。该香气物质在绿茶和红茶中主要贡献类似水果和蜂蜜的香气特征,浓度很低,但气味感知阈值极低(0.004 μg/L),在一些成品茶和茶饮料的风味中发挥重要作用[61-63]。在新鲜采摘的茶树叶片中,大马士酮基本检测不到(低于检出限),而在成品茶和茶饮品(尤其罐装或瓶装茶饮料)中可检测到一定的量[44]。已有研究表明,在红茶和绿茶加工过程中,类胡萝卜素含量呈降低趋势[34, 42, 64],其原因可能是经过加工过程的系列反应,茶鲜叶中的类胡萝卜素降解成大马士酮等香气物质[61-62, 65-66]。类胡萝卜素降解形成大马士酮存在两种方式,一是酶促反应,首先C-9和C-10双键之间的双加氧酶裂解新黄质,得到蚱蜢酮,这种酮经酶转化为联烯三醇(即大马士酮的前体物质);然后联烯三醇经酸性催化发生脱水反应生成大马士酮。二是非酶促反应,大马士酮可以直接来源于新黄质的非酶促反应降解,如茶叶加工过程中可能存在的热降解或氧化、酸性条件等均能促使该非酶促反应的发生[67]。从成品茶制成茶饮品过程中,大马士酮含量会显著增加。例如,Schuh等[62]对红茶干茶和热水浸提液的香气成分进行比较分析,结果表明即使在95℃热水中浸泡相对较短时间(150 s),大马士酮含量也能提高约30%。相比之下,在罐装或瓶装茶饮料生产过程中,一般采用更高温度(120~140℃)加热数十秒至数分钟进行高温灭菌[68],此过程会促使大马士酮含量显著增加[69-70],其主要原因可能是大马士酮的糖苷结合态前体物质发生水解反应形成大马士酮[72],该反应需要在特定条件下发生。在体外单体模拟实验中,4种糖苷结合态前体物质在强酸性(pH 2.0)和高温(90℃)下可以水解形成大马士酮,而在微酸性(pH 5.4)条件下则不能[71]。Kinoshita等[71]模拟生产罐装或瓶装绿茶饮料条件下(pH 5.4、120℃)处理10 min,却在绿茶提取液中检测到大马士酮生成;当添加4种大马士酮的糖苷结合态前体物质于绿茶提取液,相同条件处理会产生更多大马士酮;当无绿茶提取液存在时,4种前体物质进行相同处理,未检测到大马士酮。由此推测,在罐装或瓶装茶饮料生产过程中,除高温和pH外,茶饮料中其他存在的未知物质可能也会影响大马士酮的糖苷结合态前体物质水解形成大马士酮,该推测需更多实验证实[44]。综上研究表明,类胡萝卜素降解形成大马士酮物质主要发生在茶叶加工和茶饮料生产过程,且糖苷体结合的大马士酮前体物质非酶促水解是其中涉及的重要反应(图 1)。
目前β- 胡萝卜素在茶叶加工过程中降解形成香气已基本清楚,由于茶叶的加工种类和程序较复杂,下一步研究可针对茶叶干燥和冲泡的温度和时间条件进行模拟,探明茶叶中几种主要类胡萝卜素(如叶黄质、新黄质、α- 胡萝卜素和堇菜黄素)转换成的香气物质,明确各种胡萝卜素在茶叶高温加工过程中的降解产物,可为进一步改进茶叶加工工艺及饮用方式、提升茶叶品质提供理论基础。
3 展望 3.1 类胡萝卜素在茶树中的生物学功能类胡萝卜素作为代谢物在植物中具有重要生物功能,是植物重要激素ABA和独脚金内酯的前体物质。大量研究表明,ABA可参与植物生长发育和逆境胁迫响应的重要调控过程[72]。在小麦中发现高ABA含量的品种抗旱能力更强[73]。在茶树中,是否高类胡萝卜素含量对应高ABA含量目前还未明确,下一步研究可对茶树资源进行类胡萝卜素和ABA相关性进行筛查,并观察其抗胁迫能力。另外,类胡萝卜素含量不同的茶树品种在应对高温、干旱、低温及病虫害胁迫时,响应程度是否存在差异,有待进一步深入研究。
3.2 茶叶类胡萝卜素合成与代谢关键基因功能验证及调控研究茶叶中关于类胡萝卜素的研究主要集中在含量及组分分析,以及不同品种间的差异或不同处理后的变化方面。在分子机理方面,主要采用转录组和实时定量PCR的方法对茶叶中类胡萝卜素合成及代谢同源基因的表达水平进行分析。目前对茶叶类胡萝卜素关键基因的功能验证及调控机制研究还比较薄弱。虽然茶叶中的类胡萝卜素合成及代谢相关基因(CsLCYB、CsLCYE、CsCCD1和CsCCD4)已在类胡萝卜素工程菌中进行了功能验证[54-55, 74],但这些基因的体内功能和类胡萝卜素合成代谢途径的其他基因功能仍需进一步明确,此外结构基因的上游调控因子也有待进一步发掘与鉴定。这些信息将为茶树种质资源创制和茶叶加工品质调控提供必要的理论基础。
3.3 茶叶类胡萝卜素与其他代谢物合成代谢途径的交互关系越来越多研究表明,植物中的各代谢途径并不是孤立存在的,而是存在交互作用。例如,遮阴处理后茶叶呈现出高色素物质(叶绿素和类胡萝卜素)和高游离氨基酸的积累现象[75];黄化或白化的茶树品种尽管叶绿体结构不完整,但含有较高的氨基酸[76]。根据表型及生理数据推测在茶叶中质体的发育或变化往往与色素积累、氨基酸及蛋白含量呈高度相关性,但其内在的精确调控及分流机制仍需进一步研究。
总体而言,目前茶叶中类胡萝卜素的研究在生理或生化水平方面已取得较好进展,然而在分子调控机制方面的研究仍需进一步加强。此外,类胡萝卜素在茶叶采前阶段、采后加工阶段、深加工阶段和冲泡过程的动态变化与转化规律需进一步研究,从而为茶叶品质的精准调控和定向改良提供理论参数。
| [1] |
SIEFERMANN-HARMS D. Carotenoids in photosynthesis. I. Location in photosynthetic membranes and light-harvesting function[J]. Biochimica et Biophysica Acta(BBA)- Reviews on Bioenergetics, 1985, 811(4): 325-355. DOI:10.1016/0304-4173(85)90006-0 |
| [2] |
MA Y Z, HOLT N E, LI X P, NIYOGI K K, FLEMING G R. Evidence for direct carotenoid involvement in the regulation of photosynthetic light harvesting[J]. Proceedings of the National Academy of Sciences of the United States of America, 2003, 100(8): 4377-4382. DOI:10.1073/pnas.0736959100 |
| [3] |
VISHNEVETSKY M, OVADIS M, VAINSTEIN A. Carotenoid sequestration in plants: the role of carotenoid-associated proteins[J]. Trends in Plant Science, 1999, 4(6): 232-235. DOI:10.1016/S1360-1385(99)01414-4 |
| [4] |
DELLAPENNA D. Carotenoid synthesis and function in plants: Insights from mutant studies in Arabidopsis thaliana//. FRANK H A, YOUNG A J, BRITTON G, COGDELL R J. The photochemistry of carotenoids[M]. Dordrecht; Springer Netherlands, 1999. DOI: 10.1007/0-306-48209-62.
|
| [5] |
LUAN Y, FU X, LU P, GRIERSON D, XU C. Molecular mechanisms determining the differential accumulation of carotenoids in plant species and varieties[J]. Critical Reviews in Plant Sciences, 2020, 39(2): 125-139. DOI:10.1080/07352689.2020.1768350 |
| [6] |
LEWINSOHN E, SITRIT Y, BAR E, AZULAY Y, MEIR A, ZAMIR D, TADMOR Y. Carotenoid pigmentation affects the volatile composition of tomato and watermelon fruits, as revealed by comparative genetic analyses[J]. Journal of Agricultural and Food Chemistry, 2005, 53(8): 3142-3148. DOI:10.1021/jf047927t |
| [7] |
LEFEBVRE V, KUNTZ M, CAMARA B, PALLOIX A. The capsanthin-capsorubin synthase gene: a candidate gene for the y locus controlling the red fruit colour in pepper[J]. Plant Molecular Biology, 1998, 36(5): 785-789. DOI:10.1023/A:1005966313415 |
| [8] |
FU X, KONG W, PENG G, ZHOU J, AZAM M, XU C, GRIERSON D, CHEN K. Plastid structure and carotenogenic gene expression in red-and white-fleshed loquat(Eriobotrya japonica)fruits[J]. Journal of Experimental Botany, 2011, 63(1): 341-354. DOI:10.1093/jxb/err284 |
| [9] |
XU C J, FRASER P D, WANG W J, BRAMLEY P M. Differences in the carotenoid content of ordinary citrus and lycopene-accumulating mutants[J]. Journal of Agricultural and Food Chemistry, 2006, 54(15): 5474-5481. DOI:10.1021/jf060702t |
| [10] |
LIU Y, ZENG S, SUN W, WU M, HU W, SHEN X, WANG Y. Comparative analysis of carotenoid accumulation in two goji(Lycium barbarum L. and L. ruthenicum Murr.)fruits[J]. BMC Plant Biology, 2014, 14(1): 269. DOI:10.1186/s12870-014-0269-4 |
| [11] |
NGAMWONGLUMLERT L, DEVAHASTIN S, CHIEWCHAN N, RAGHAVAN V. Plant carotenoids evolution during cultivation, postharvest storage, and food processing: A review[J]. Comprehensive Reviews in Food Science and Food Safety, 2020, 19(4): 1561-1604. DOI:10.1111/1541-4337.12564 |
| [12] |
曾晓雄. 茶叶中类胡萝卜素的氧化降解及其与茶叶品质的关系[J]. 茶叶通讯, 1992(1): 31-33. ZENG X X. Oxidative degradation of carotenoids in tea and its relationship with tea quality[J]. Journal of Tea Communication, 1992(1): 31-33. |
| [13] |
宛晓春, 夏涛. 茶树次生代谢[M]. 北京: 科学出版社, 2015. WAN X C, XIA T. The secondary metabolism of tea tree[M]. Beijing: Science Press, 2015. |
| [14] |
王松琳, 马春雷, 黄丹娟, 马建强, 金基强, 姚明哲, 陈亮. 基于SSR标记的白化和黄化茶树品种遗传多样性分析及指纹图谱构建[J]. 茶叶科学, 2018, 38(1): 58-68. DOI:10.13305/j.cnki.jts.2018.01.006 WANG S L, MA C L, HUANG D J, MA J Q, JIN J Q, YAO M Z, CHEN L. Analysis of genetic diversity and construction of DNA fingerprints of chlorophyll-deficient tea cultivars by SSR markers[J]. Journal of Tea Science, 2018, 38(1): 58-68. DOI:10.13305/j.cnki.jts.2018.01.006 |
| [15] |
卢翠, 沈程文. 茶树白化变异研究进展[J]. 茶叶科学, 2016, 36(5): 445-541. DOI:10.13305/j.cnki.jts.2016.05.001 LU C, SHEN C W. Research progress of albino tea plant(Camellia sinensis L. O. Kuntze)[J]. Journal of Tea Science, 2016, 36(5): 445-451. DOI:10.13305/j.cnki.jts.2016.05.001 |
| [16] |
WANG L, YUE C, CAO H, ZHOU Y, ZENG J, YANG Y, WANG X. Biochemical and transcriptome analyses of a novel chlorophyll-deficient chlorina tea plant cultivar[J]. BMC Plant Biology, 2014, 14(1): 352. DOI:10.1186/s12870-014-0352-x |
| [17] |
FENG L, GAO M J, HOU R Y, HU X Y, ZHANG L, WAN X C, WEI S. Determination of quality constituents in the young leaves of albino tea cultivars[J]. Food Chemistry, 2014, 155: 98-104. DOI:10.1016/j.foodchem.2014.01.044 |
| [18] |
李明, 张龙杰, 石萌, 林小明, 王开荣, 郑新强, 陆建良, 梁月荣. 光照敏感型新梢白化茶新品种'御金香'春梢化学成分研究[J]. 茶叶, 2016, 42(3): 146-149. DOI:10.3969/j.issn.0577-8921.2016.03.004 LI M, ZHANG L J, SHI M, LIN X M, WANG K R, ZHENG X Q, LU J L, LIANG Y R. Chemical composition of spring shoots in light-sensitive albino tea cultivar'Yujinxiang'[J]. Journal of Tea, 2016, 42(3): 146-149. DOI:10.3969/j.issn.0577-8921.2016.03.004 |
| [19] |
ZHAO C, SAFDAR L B, XIE M, SHI M, DONG Z, YANG L, CHENG X, LIU Y, BAI Z, XIANG Y, TONG C, HUANG J, LIU L, LIU S. Mutation of the PHYTOENE DESATURASE 3 gene causes yellowish-white petals in Brassica napus[J]. The Crop Journal, 2021. DOI:10.1016/j.cj.2020.10.012 |
| [20] |
李娜娜, 邵文韵, 刘畅, 陆建良, 梁月荣. 茶树八氢番茄红素脱氢酶cDNA全长克隆与表达分析[J]. 茶叶, 2014, 40(2): 69-74. DOI:10.3969/j.issn.0577-8921.2014.02.002 LI N N, SHAO W Y, LIU C, LU J L, LIANG Y R. Cloning of a full-length cDNA of phytoene desaturase gene in tea plant(Camellia sinensis)and its expression[J]. Journal of Tea, 2014, 40(2): 69-74. DOI:10.3969/j.issn.0577-8921.2014.02.002 |
| [21] |
LI C F, XU Y X, MA J Q, JIN J Q, HUANG D J, YAO M Z, MA C L, CHEN L. Biochemical and transcriptomic analyses reveal different metabolite biosynthesis profiles among three color and developmental stages in 'Anji Baicha'(Camellia sinensis)[J]. BMC Plant Biology, 2016, 16(1): 195. DOI:10.1186/s12870-016-0885-2 |
| [22] |
ZHENG Y, WANG P, CHEN X, YUE C, GUO Y, YANG J, SUN Y, YE N. Integrated transcriptomics and metabolomics provide novel insight into changes in specialized metabolites in an albino tea cultivar (Camellia sinensis L. O. Kuntz)[J]. Plant Physiology and Biochemistry, 2021, 160: 27-36. DOI:10.1016/j.plaphy.2020.12.029 |
| [23] |
MATSUBARA S, KRAUSE G H, ARANDA J, VIRGO A, BEISEL K G, JAHNS P, WINTER K. Sun-shade patterns of leaf carotenoid composition in 86 species of neotropical forest plants[J]. Functional Plant Biology, 2009, 36(1): 20-36. DOI:10.1071/FP08214 |
| [24] |
JOHNSON G N, SCHOLES J D, HORTON P, YOUNG A J. Relationships between carotenoid composition and growth habit in British plant species[J]. Plant Cell & Environment, 1993, 16(6): 681-686. DOI:10.1111/j.1365-3040.1993.tb00486.x |
| [25] |
WANG X C, CHEN L, MA C L, YAO M Z, YANG Y J. Genotypic variation of beta-carotene and lutein contents in tea germplasms, Camellia sinensis L.O.Kuntze[J]. Journal of Food Composition and Analysis, 2010, 23(1): 9-14. DOI:10.1016/j.jfca.2009.01.016 |
| [26] |
WEI X, HOU Y J, YE Y L, ZHANG Q Y, ZHENG H W, LIANG G L, TONG H R. Determination of carotenoid contents in 31 elite Chinese tea[Camellia sinensis(L.)O.Kuntze]cultivars[J]. Academia Journal of Agricultural Research, 2016, 4(8): 525-533. DOI:10.15413/ajar.2016.0173 |
| [27] |
陈丽, 坤吉瑞, 王静, 童华荣. HPLC-PDA法分析不同茶树品种类胡萝卜素香气前体[J]. 食品科学, 2020, 41(4): 193-198. DOI:10.7506/spkx1002-6630-20190318-227 CHEN L, KUN J R, WANG J, TONG H R. Analysis of carotenoids as aroma precursors in different varieties of tea leaves by high performance liquid chromatography with photodiode array detection[J]. Food Science, 2020, 41(4): 193-198. DOI:10.7506/spkx1002-6630-20190318-227 |
| [28] |
BORTHAKUR D L J, CHEN H, LIN C, DU Y Y, LIANG Y R. Expression of phytoene synthase(psy)gene and its relation with accumulation of carotenoids in tea Camellia sinensis L O Kuntze][J]. African Journal of Biotechnology, 2008, 7(4): 434-438. |
| [29] |
VENKATAKRISHNA S, PREMACHANDRA B R, CAMA H R. Comparative study of the effects of processing on the carotenoid composition of Chinese(Thea sinensis)and Assamese(Thea assamica)tea[J]. Agricultural and Biological Chemistry, 1976, 40(12): 2367-2371. DOI:10.1080/00021369.1976.10862420 |
| [30] |
RAVICHANDRAN R. Carotenoid composition, distribution and degradation to flavour volatiles during black tea manufacture and the effect of carotenoid supplementation on tea quality and aroma[J]. Food Chemistry, 2002, 78(1): 23-28. DOI:10.1016/S0308-8146(01)00303-X |
| [31] |
YOO S D, GREER D H, LAING W A, MCMANUS M T. Changes in photosynthetic efficiency and carotenoid composition in leaves of white clover at different developmental stages[J]. Plant Physiology and Biochemistry, 2003, 41(10): 887-893. DOI:10.1016/S0981-9428(03)00138-4 |
| [32] |
VENKATAKRISHNA S, PREMACHANDRA B R, CAMA H R. Distribution of carotenoid pigments in tea leaves[J]. Tea Quarterly, 1977, 47(1 & 2): 28-31. |
| [33] |
SONOBE R, MIURA Y, SANO T, HORIE H. Estimating leaf carotenoid contents of shade-grown tea using hyperspectral indices and PROSPECT–D inversion[J]. International Journal of Remote Sensing, 2018, 39(5): 1306-1320. DOI:10.1080/01431161.2017.1407050 |
| [34] |
KAWAKAM MICHIKO Y T. Aroma characteristics of Kabusecha (shaded green tea)[J]. Nippon Nogeikagaku Kaishi, 1981, 55: 117-123. DOI:10.1271/nogeikagaku1924.55.117 |
| [35] |
王加真, 张昕昱, 金星, 黄家春, 陈莹莹. 红蓝光源茶园夜间补光对春茶产量、品质的影响[J]. 福建农业学报, 2019, 34(1): 46-52. DOI:10.19303/j.issn.1008-0384.2019.01.008 WANG J Z, ZHANG X Y, JIN X, HUANG J C, CHEN Y Y. Effects of red and blue led light at night on growth and leaf quality of tea plant[J]. Fujian Journal of Agricultural Sciences, 2019, 34(1): 46-52. DOI:10.19303/j.issn.1008-0384.2019.01.008 |
| [36] |
范延艮'. 黄金芽'茶树不同色泽新梢多组学比较及生理特性研究[D]. 泰安: 山东农业大学, 2019. DOI: 10.27277/d.cnki.gsdnu.2019.000018. FAN Y G. Multiomics comparison and physiological characteristics of different colour shoots of Camellia sinensis var. Huangjingya[D]. Taian: Shandong Agricultural University, 2019. DOI: 10.27277/d.cnki.gsdnu.2019.000018.gsdnu.2019.000018. |
| [37] |
SONG L, MA Q, ZOU Z, SUN K, YAO Y, TAO J, KALERI N A, LI X. Molecular link between leaf coloration and gene expression of flavonoid and carotenoid biosynthesis in Camellia sinensis cultivar 'Huangjinya'[J]. Frontiers in Plant Science, 2017, 8: 803. DOI:10.3389/fpls.2017.00803 |
| [38] |
FU X, CHENG S, FENG C, KANG M, HUANG B, JIANG Y, DUAN X, GRIERSON D, YANG Z. Lycopene cyclases determine high α-/β-carotene ratio and increased carotenoids in bananas ripening at high temperatures[J]. Food Chemistry, 2019, 283: 131-140. DOI:10.1016/j.foodchem.2018.12.121 |
| [39] |
DU Y Y, CHEN H, ZHONG W L, WU L Y, YE J H, LIN C, ZHENG X Q, LU J L, LIANG Y R. Effect of temperature on accumulation of chlorophylls and leaf ultrastructure of low temperature induced albino tea plant[J]. African Journal of Biotechnology, 2008, 7(12): 1881-1885. DOI:10.5897/AJB2008.000-5036 |
| [40] |
DU Y Y, SHIN S, WANG K R, LU J L, LIANG Y R. Effect of temperature on the expression of genes related to the accumulation of chlorophylls and carotenoids in albino tea[J]. The Journal of Horticultural Science and Biotechnology, 2009, 84(3): 365-369. DOI:10.1080/14620316.2009.11512533 |
| [41] |
邓婷婷. 不同温度下安吉白茶差异表达基因的分离、鉴定及CsPDS3基因的功能研究[D]. 长沙: 湖南农业大学, 2013. DENG T T. Isolation and identification of differential expressed genes under different temperatures from Anji Baicha[Camellia sinensis L.O. Kuntze] and functional analysis of CsPDS3 gene[D]. Changsha: Hunan Agricultural University, 2013. |
| [42] |
HAZARIKA M, MAHANTA P K. Some studies on carotenoids and their degradation in black tea manufacture[J]. Journal of the Science of Food and Agriculture, 1983, 34(12): 1390-1396. DOI:10.1002/jsfa.2740341212 |
| [43] |
AULDRIDGE M E, MCCARTY D R, KLEE H J. Plant carotenoid cleavage oxygenases and their apocarotenoid products[J]. Current Opinion in Plant Biology, 2006, 9(3): 315-321. DOI:10.1016/j.pbi.2006.03.005 |
| [44] |
YANG Z, BALDERMANN S, WATANABE N. Chapter 31 - Formation of damascenone and its related compounds from carotenoids in tea//PREEDY V R. Tea in health and disease prevention[M]. Academic Press. 2013: 375-386. DOI: 10.1016/B978-0-12-384937-3.00031-8.
|
| [45] |
陈丽, 叶玉龙, 王春燕, 闫敬娜, 童华荣. 茶叶中类胡萝卜素香气前体研究进展[J]. 食品与发酵工业, 2019, 45(5): 266-273. DOI:10.13995/j.cnki.11-1802/ts.017973.ts.017973 CHEN L, YE Y L, WANG C Y, YAN J N, TONG H R. Research progress on carotenoid aroma precursors in tea[J]. Food and Fermentation Industries, 2019, 45(5): 266-273. DOI:10.13995/j.cnki.11-1802/ts.017973.ts.017973 |
| [46] |
刘飞, 王云, 张厅, 唐晓波, 王小萍, 李春华. 红茶加工过程香气变化研究进展[J]. 茶叶科学, 2018, 38(1): 9-19. DOI:10.13305/j.cnki.jts.2018.01.002 LIU F, WANG Y, ZHANG T, TANG X B, WANG X P, LI C H. Review on aroma change during black tea processing[J]. Journal of Tea Science, 2018, 38(1): 9-19. DOI:10.13305/j.cnki.jts.2018.01.002 |
| [47] |
SANDERSON G W, GRAHAMM H N. Formation of black tea aroma[J]. Journal of Agricultural and Food Chemistry, 1973, 21(4): 576-585. DOI:10.1021/jf60188a007 |
| [48] |
FLEISCHMANN P, BALDERMANN S, YAMAMOTO M, WATANABE N, WINTERHALTER P. Identification, isolation, and evaluation of carotenoid degradation enzymes in japanese green tea(Camellia sinensis, THEACEAE)[A]. Proceedings of 2005 international symposium on innovation in tea science and sustainable development in tea industry[C]. 2005.
|
| [49] |
FELFE C, SCHEMAINDA M, BALDERMANN S, WATANABE N, FLEISCHMANN P. Metabolism of carotenoid degradation in leaves of Camellia sinensis—Functional and biochemical modifications[J]. Journal of Food Composition and Analysis, 2011, 24(6): 821-825. DOI:10.1016/j.jfca.2011.03.017 |
| [50] |
FLOSS D S, WALTER M H. Role of carotenoid cleavage dioxygenase 1 (CCD1)in apocarotenoid biogenesis revisited[J]. Plant Signaling & Behavior, 2009, 4(3): 172-175. DOI:10.4161/psb.4.3.7840 |
| [51] |
OHMIYA A, KISHIMOTO S, AIDA R, YOSHIOKA S, SUMITOMO K. Carotenoid cleavage dioxygenase(CmCCD4a)contributes to white color formation in Chrysanthemum petals[J]. Plant Physiology, 2006, 142(3): 1193-1201. DOI:10.1104/pp.106.087130 |
| [52] |
HUANG F C, MOLNÁR P, SCHWAB W. Cloning and functional characterization of carotenoid cleavage dioxygenase 4 genes[J]. Journal of Experimental Botany, 2009, 60(11): 3011-3022. DOI:10.1093/jxb/erp137 |
| [53] |
王赞, 岳川, 曹红利, 郭雅玲. 茶树CsCCD1和CsCCD4基因的克隆和表达分析[J]. 西北植物学报, 2018, 38(9): 1605-1612. WANG Z, YUE C, CAO H L, GUO Y L. Cloning and expression analysis of CsCCD1and CsCCD4 in tea plant(Camellia sinensis)[J]. Acta Botanica Boreali-Occidentalia Sinca, 2018, 38(9): 1605-1612. |
| [54] |
WANG J, ZHANG N, ZHAO M, JING T, JIN J, WU B, WAN X, SCHWAB W, SONG C. Carotenoid cleavage dioxygenase 4 catalyzes the formation of carotenoid-derived volatile β-ionone during tea (Camellia sinensis)withering[J]. Journal of Agricultural and Food Chemistry, 2020, 68(6): 1684-1690. DOI:10.1021/acs.jafc.9b07578 |
| [55] |
WANG J, WU B, ZHANG N, ZHAO M, JING T, WU Y, HU Y, YU F, WAN X, SCHWAB W, SONG C. Dehydration-induced carotenoid cleavage dioxygenase 1 reveals a novel route for β-ionone formation during tea (Camellia sinensis)withering[J]. Journal of Agricultural and Food Chemistry, 2020, 68(39): 10815-10821. DOI:10.1021/acs.jafc.0c04208 |
| [56] |
BOKUCHAVA M A, SKOBELEVA N I. The chemistry and biochemistry of tea and tea manufacture//CHICHESTER C O, MRAK E M, STEWART G F. Advances in food research[M]. Academic Press, 1969. DOI: 10.1016/S0065-2628(08)60311-0.
|
| [57] |
川 嶋か, 山西貞. Beta-carotene の熱分解[J]. 日本農芸化学会誌, 1973, 47(1): 79-81. DOI:10.1271/nogeikagaku1924.47.79 KAWASHIMA K, YAMANISHI T. Thermal-degradation of beta-carotene[J]. Journal of Japan Society for Bioscience and Biotechnology, 1973, 47(1): 79-81. DOI:10.1271/nogeikagaku1924.47.79 |
| [58] |
KANASAWUD P, CROUZET J C. Mechanism of formation of volatile compounds by thermal degradation of carotenoids in aqueous medium. 1.β-Carotene degradation[J]. Journal of Agricultural and Food Chemistry, 1990, 38(1): 237-243. DOI:10.1021/jf00091a052 |
| [59] |
吕世懂, 吴远双, 姜玉芳, 孟庆雄. 不同产区乌龙茶香气特征及差异分析[J]. 食品科学, 2014, 35(2): 146-153. LYU S D, WU Y S, JIANG Y F, MENG Q X. Comparative analysis of aroma characteristics of oolong tea from different geographical regions[J]. Food Science, 2014, 35(2): 146-153. |
| [60] |
SEFTON M A, SKOUROUMOUNIS G K, ELSEY G M, TAYLOR D K. Occurrence, sensory impact, formation, and fate of damascenone in grapes, wines, and other foods and beverages[J]. Journal of Agricultural and Food Chemistry, 2011, 59(18): 9717-9746. DOI:10.1021/jf201450q |
| [61] |
KUMAZAWA K, MASUDA H. Identification of potent odorants in different green tea varieties using flavor dilution technique[J]. Journal of Agricultural and Food Chemistry, 2002, 50(20): 5660-5663. DOI:10.1021/jf020498j |
| [62] |
SCHUH C, SCHIEBERLE P. Characterization of the key aroma compounds in the beverage prepared from darjeeling black tea: Quantitative differences between tea leaves and infusion[J]. Journal of Agricultural and Food Chemistry, 2006, 54(3): 916-924. DOI:10.1021/jf052495n |
| [63] |
PRIPDEEVECH P, MACHAN T. Fingerprint of volatile flavour constituents and antioxidant activities of teas from Thailand[J]. Food Chemistr y, 2011, 125(2): 797-802. DOI:10.1016/j.foodchem.2010.09.074 |
| [64] |
SANDERSON G W, CO H, GONZALEZ J G. Biochemistry of tea fermentation: The role of carotenes in black tea aroma formation[J]. Journal of Food Science, 1971, 36(2): 231-236. DOI:10.1111/j.1365-2621.1971.tb04031.x |
| [65] |
KAWAKAMI M, GANGULY S N, BANERJEE J, KOBAYASHI A. Aroma composition of oolong tea and black tea by brewed extraction method and characterizing compounds of darjeeling tea aroma[J]. Journal of Agricultural and Food Chemistry, 1995, 43(1): 200-207. DOI:10.1021/jf00049a037 |
| [66] |
KAWAKAMI M, KOBAYASHI A. Carotenoid-derived aroma compounds in tea. Carotenoid-derived aroma compounds[M]. American Chemical Society, 2001. DOI: 10.1021/bk-2002-0802.ch011.
|
| [67] |
HO C T, ZHENG X, LI S. Tea aroma formation[J]. Food Science and Human Wellness, 2015, 4(1): 9-27. DOI:10.1016/j.fshw.2015.04.001 |
| [68] |
MASUDA H. Flavor stability of tea drinks. HO C T, LIN J K, SHAHIDI, F. Tea and tea products: Chemistry and health-promoting properties[M]. CRC Press Taylor & Francis Group: Boca Raton. 2008. DOI: 10.1201/9781420008036.
|
| [69] |
KUMAZAWA K, MASUDA H. Change in the flavor of black tea drink during heat processing[J]. Journal of Agricultural and Food Chemistry, 2001, 49(7): 3304-3309. DOI:10.1021/jf001323h |
| [70] |
KUMAZAWA K. Flavor chemistry of tea and coffee drinks[J]. Food Science Technology Research, 2006, 12: 71-84. DOI:10.3136/fstr.12.71 |
| [71] |
KINOSHITA T, HIRATA S, YANG Z, BALDERMANN S, KITAYAMA E, MATSUMOTO S, SUZUKI M, FLEISCHMANN P, WINTERHALTER P, WATANABE N. Formation of damascenone derived from glycosidically bound precursors in green tea infusions[J]. Food Chemistry, 2010, 123(3): 601-606. DOI:10.1016/j.foodchem.2010.04.077 |
| [72] |
BULGAKOV V P, WU H C, JINN T L. Coordination of ABA and chaperone signaling in plant stress responses[J]. Trends in Plant Science, 2019, 24(7): 636-651. DOI:10.1016/j.tplants.2019.04.004 |
| [73] |
康书江, 赵春江, 杨宝祝. 小麦内源激素脱落酸研究进展[J]. 麦类作物学报, 1997(3): 47-51. KANG S J, ZHAO C J, YANG B Z. Research progress of wheat endogenous hormone abscisic acid[J]. Tritical Crops, 1997(3): 47-51. |
| [74] |
刘关华, 杨梅, 付建玉. 茶树胡萝卜素合成关键基因CsLCYb和CsLCYe的克隆与功能鉴定[J]. 茶叶科学, 2019, 39(3): 257-266. DOI:10.13305/j.cnki.jts.2019.03.003.j.cnki.jts.2019.03.003 LIU G H, YANG M, FU J Y. Cloning and functional analysis of CsLCYb and CsLCYe for carotene biosynthesis in tea plant(Camellia sinensis)[J]. Journal of Tea Science, 2019, 39(3): 257-266. DOI:10.13305/j.cnki.jts.2019.03.003.j.cnki.jts.2019.03.003 |
| [75] |
YU Z, YANG Z. Understanding different regulatory mechanisms of proteinaceous and non-proteinaceous amino acid formation in tea(Camellia sinensis)provides new insights into the safe and effective alteration of tea flavor and function[J]. Critical Reviews in Food Science and Nutrition, 2020, 60(5): 844-858. DOI:10.1080/10408398.2018.1552245 |
| [76] |
LU M, HAN J, ZHU B, JIA H, YANG T, WANG R, DENG W W, ZHANG Z Z. Significantly increased amino acid accumulation in a novel albino branch of the tea plant(Camellia sinensis)[J]. Planta, 2019, 249(2): 363-376. DOI:10.1007/s00425-018-3007-6 |