 2022, Vol. 49
2022, Vol. 49文章信息
引用本文 |

基金项目
- 国家自然科学基金(31870380)
作者简介
- 宋晨慧(1997—),女,在读硕士生,研究方向为植物生理生态,E-mail:sch6057@163.com.
通讯作者
- 张有福(1978—),男,博士,副教授,研究方向为木本植物生理生态和环境地球化学,E-mail:zyfgau@163.com.
文章历史
- 收稿日期:2022-02-12

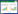


【研究意义】植物气孔是由两个特殊分化的保卫细胞构成的孔状结构[1],是植物与环境进行气体和水分交换的通道,也是植物响应环境变化调节光合和蒸腾的调节位点。为适应复杂多变的环境,植物既可通过保卫细胞的膨压调节气孔大小和开度,也可通过气孔密度和气孔分布平衡光合作用和水分散失[2]。其中植物依赖于气孔的开度调节蒸腾和光合常用来解释适应环境因子的短期变化[3],气孔分布和密度的变化则是植物适应环境因子长期变化的结果。然而植物往往通过多种气孔功能性状协调适应环境变化,表现出气孔大小与气孔密度之间存在稳定的负相关关系[4]。另外,气孔特征也受到植物本身遗传因素的决定。因此,分析不同环境气孔特征、气孔参数之间的关系有利于揭示植物适应环境的种间差异和长期与短期适应的差异。【前人研究进展】光是植物赖以生存的基础。光照强度被认为是影响植物气孔发育和功能的重要环境因素。长期以来,关于气孔性状对光照强度的响应展开了大量的研究并取得一些成果。花生通过保卫细胞的膨胀和收缩调节气孔大小以适应光照的日变化[5]。10种地被植物在遮荫环境下叶片气孔密度相对较少,小的气孔密度有利于叶片更好地吸收并储存光能[6]。枫香(Liquidambar formosana)通过调节气孔开度和气孔密度以适应不同环境的长期光照差异[7]。蛇足石杉(Huperzia serrata)在一定范围内,随着环境中光照强度的增加,气孔长度和宽度、气孔密度都呈增加的趋势,小而密的气孔是其适应旱生环境的表现[8]。刺槐(Robinia pseudoacacia)在光照充足的南坡叶片气孔小但气孔密度高,在郁闭程度高的北坡气孔大但气孔密度小[9],这是光合产物在不同构件或功能间权衡分配的结果。
【本研究切入点】以往的研究倾向于气孔特征的日变化来解释植物适应短期光照变化下的气孔调节,或者不同生境下气孔差异解析长期光照差异下气孔的适应性变化[5, 7],但这些研究都缺乏气孔结构和功能在同一环境下长短期光照变化的比较分析。因此,比较同一生境下同属植物气孔响应长期和短期光照差异将为进一步阐明植物气孔响应光环境变化的适应机制提供借鉴。【拟解决的关键问题】本研究选取3种圆柏属植物为研究对象,测定同一生境不同光照下的气孔特征并进行比较分析,进一步探讨气孔响应光环境变化及其适应策略,为解析圆柏属植物叶形差异与光照强度关系与适应性提供理论依据。
1 材料与方法 1.1 试验材料供试样品采自洛阳市周山森林公园,该区属暖温带大陆性季风气候,海拔196~269 m,年均降水量672.6 mm。2021年3月中旬,在地势平坦、树间无明显遮荫的地方,选择生长情况良好、健壮无虫害的圆柏(Juniperus chinensis)、龙柏(J. chinensis cv. Kaizuca)、塔柏(J. chinensis cv. Pyramidalis)各4株,选择无风、晴朗的天气,剪取离地2 m处的树冠阴、阳面的叶片,将其分别放入装有FAA固定液的离心管中,带回实验室抽真空至叶片全部沉入固定液中,24 h后进行制片和气孔特征测定。因圆柏蒸腾速率的日变化曲线基本呈单峰抛物线型,13: 00左右达到最高峰[10]。采样时间为早上5: 50—6: 20和中午11: 00—11: 30,即此时间段内,光照为影响气孔运动的主要因素。
3种圆柏属植物均具异形叶特征,其中圆柏、龙柏多鳞形叶,塔柏多刺形叶。因此,在进行叶片气孔特征比较时,为避免叶型带来的影响,圆柏和龙柏的叶片仅采用鳞形叶,塔柏仅采用刺形叶。以早晨和中午时段采集的叶片代表短期光照的变化;因树冠朝向导致的叶片所处的光环境不同,以树冠阴、阳面代表长期光照差异。
1.2 试验方法叶表皮制片方法参照朱玉怀等的NaClO法[11]。每个树种选取8张临时装片用于测定各气孔参数,每张制片在显微镜下随机选取5个视野,每个视野内随机选取5个气孔,用显微测微尺测量保卫细胞长度(GCL)和保卫细胞宽度(GCW)及气孔开度(SA),气孔开度由气孔内壁最宽处的宽度代表,并记录视野面积内的气孔个数,计算气孔密度(SD):
气孔密度=气孔数/视野面积(0.1590 mm2)
自然界中多数植物的气孔主要分布在叶片远轴面,但鉴于圆柏属植物的气孔主要分布在叶片近轴面,故仅以近轴面气孔作相关分析。
1.3 数据分析对不同自然光强下植物叶片的气孔参数进行配对样本t检验。若差异显著,则将显著变化的气孔参数组合,采用线性回归法分析它们在长短期光照变化下的关系。采用IBM SPSS Statistics 22.0软件进行统计分析,应用Microsoft Excel 2019软件作图。
2 结果与分析 2.1 圆柏属植物叶片气孔特征圆柏属植物存在鳞形叶和刺形叶,圆柏和龙柏鳞形叶上的气孔主要分布在近轴面上,除了中轴和叶缘处,整个近轴面都分布有气孔,而远轴面上只有相邻两叶交界处有气孔分布。塔柏刺形叶的气孔仅分布在叶片近轴面,除了中轴处和叶缘外,整个近轴面全部被气孔覆盖,且有两条明显的气孔带,而远轴面无气孔分布(表 1)。可见,具有异形叶的圆柏属植物的气孔多分布在叶的近轴面。
|
2.2 自然光照对3种圆柏属植物气孔参数的影响
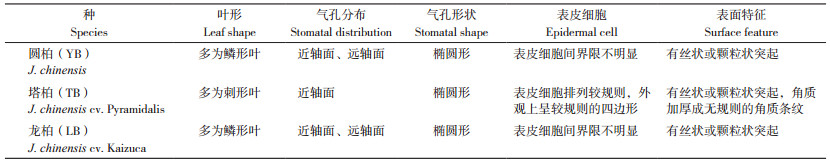
2.2.1 短期光照变化对气孔参数的影响 3种圆柏属植物气孔参数的日变化趋势基本一致(表 2)。中午自然光照强度增强时,3种圆柏属植物GCW比早晨分别显著减小0.06%、0.09%、0.09%,SA分别显著增大27.6%、24.3%、24.3%,但GCL、SD无显著差异,表明3种圆柏属植物主要通过调节GCW的收缩增大SA以适应短期光照变化。
|
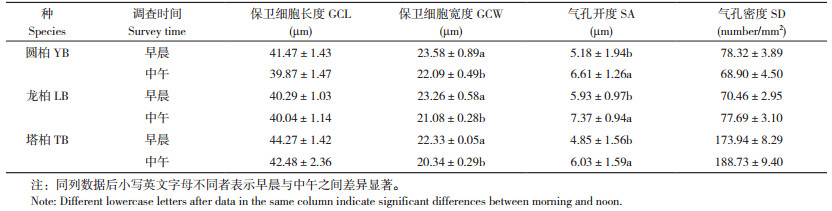
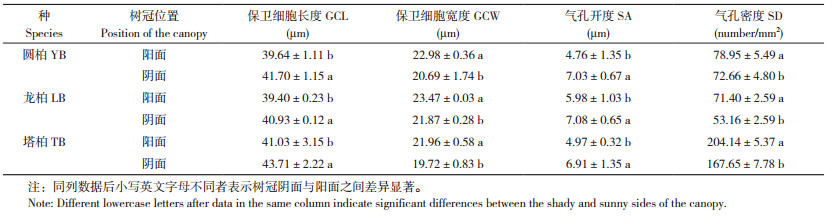
2.2.2 长期光照差异对气孔参数的影响 树冠朝向引起叶片接受的光照强弱和时长不同,导致3种圆柏属植物阴阳面气孔参数间存在差异(表 3)。圆柏、龙柏、塔柏阴面叶的GCL比阳面分别显著增大0.05%、0.04%、0.07%,SA比阳面分别显著增大47.7%、18.4%、39.0%,但GCW比阳面分别显著减小1.0%、6.8%、10.2%,SD比阳面分别显著降低26%、8%、18%,表明植物通过SA和SD的双重调节以适应长期光照差异。
|
2.3 自然光照下3种圆柏属植物气孔参数间的关系
2.3.1 短期光照变化下气孔参数间的关系 日变化下,3种圆柏属植物叶片的GCW与SA均发生显著变化且相关性较强(r分别为0.879、0.914、0.418)(图 1),表明3种圆柏属植物气孔参数相关性一致,SA均随着GCW的减小而增大。

|
| 图 1 3种圆柏属植物日变化下气孔参数间的相关性 Fig. 1 Correlation of stomatal parameters of three species of Juniperus under diurnal changes |
2.3.2 长期光照差异下气孔参数间的关系 3种圆柏属植物叶片的GCL、GCW、SA、SD在树冠阴阳面都有显著差异(图 2)。将SA与GCL、GCW、SD组合进行线性回归分析表明,3种圆柏属植物叶片的SA与GCL均呈正相关关系(r分别为0.880、0.752、0.918),与GCW均呈负相关关系(r分别为0.891、0.857、0.818),与SD也均呈负相关关系分别为(r分别为0.898、0.937、0.500)(图 2)。可见,SA随着GCL的增加而增大,随着GCW、SD的增大而减小。

|
| 图 2 3种圆柏属植物树冠阴阳面气孔参数间的相关性 Fig. 2 Correlation of stomatal parameters between the shady and sunny sides of the canopy of three species of Juniperus |
3 讨论 3.1 圆柏属植物异形叶间气孔特征的差异
圆柏属植物有鳞形和刺形两种叶形。本研究前期工作发现,鳞形叶紧贴小枝,气孔分布在近轴面以及两鳞形叶交界处的远轴面上;刺形叶与小枝分离,气孔仅分布于近轴面,有两条明显的气孔带。这与邵邻相等[12]对柏科植物叶片气孔分布的研究结果一致。为了保证积累足够的营养物质,以求得生存与繁衍,植物在长期进化过程一定会保留有利于保持旺盛光合、呼吸代谢活力的气孔特征[13]。圆柏属植物的叶形狭小,远轴面比近轴面更多地暴露于空气中和阳光下,因此气孔倾向于分布在相对远离外界环境,受到相对“保护”的近轴面,这种分布有效抑制了叶内的蒸腾,降低了水分的蒸发,提高了叶子的抗早性。这与吕洪飞[13]对双子叶植物的研究结果不一致,青藏高原双子叶植物气孔常分布在远轴面,这种分布方式有利于其与外界环境进行二氧化碳和水分的交换,同时适当的抑制蒸腾作用,避免水分过度蒸发而造成生理干旱,这是双子叶植物对高寒极端环境长期适应的结果。可见,气孔分布与叶形的关系及其差异在一定程度上反映了植物对环境长期适应。
3.2 气孔特征对短期光照变化的响应依赖于气孔开度调节的气孔运动被认为是植物适应环境变化重要反应[14]。本研究发现,在日变化过程中,植物通过降低保卫细胞宽度以增加气孔开度来适应短期光照变化。这与王萍等[15]研究结果一致,日变化过程中气孔开度主要依赖于保卫细胞宽度的收缩。由于圆柏蒸腾速率的日变化曲线均呈单峰抛物线型,峰值出现在13:00左右[10],因此,本研究中,与早晨相比,中午光强增加时,气孔开度的增加能够提高植物叶片光合作用和蒸腾作用,同时降低叶温,促进水分及无机盐的吸收和运输[15]。此外,当气孔发育过程中保卫细胞宽度生长速率大于保卫细胞长度生长速率时,气孔会逐渐向“圆”形发展,增加了单位周长下的气孔面积,这有助于减小叶表气孔器的物质与能量投资[16]。因此,我们推断植物主要采用低耗能的保卫细胞宽度改变来调节气孔开度以适应短期光照变化。
3.3 气孔特征对长期光照差异的响应树冠朝向不同,叶片接受的光照长期存在差异,可用于探索叶片对长期光照环境变化的适应差异。本研究中,长期光照差异下,通过保卫细胞长度和宽度双重调节气孔的开度,与气孔密度在叶片生长发育过程中的变化是植物对长期光照差异适应的方式。这种调节方式与其他物种的研究结果基本一致[17]。气孔直接影响植物的CO2交换速率、蒸腾速率和光合速率,不同光照下气孔开度和气孔密度的组合调节被认为是植物平衡水碳循环的一种经济方式[18]。与树冠阳面不同,树冠阴面光资源不足,大气孔开度和小气孔密度更具有适应意义。植物通过增大气孔开度,或者形成“无光合午休”等机制提高CO2固定效率,在获得最大CO2的同时,还能够对气孔密度的减小具有一定的补偿作用,有利于植物在低光照条件下提高光能利用效率[19]。虽然较大的气孔开度可以交换更多的碳,但这种交换存在风险。随着气孔开度逐渐增大,其进一步开张的阻力也越大、耗费能量越多,它们的慢动力学特性也使大气孔植物在遭遇干旱等环境胁迫时更容易发生功能紊乱[20-21]。树冠阳面叶片的气孔开度小且气孔密度高,高气孔密度有利于增强叶片气体交换的能力,这样当部分气孔生理活动受到抑制时,其余功能正常的气孔能做出适度调节,在功能上及时补偿,使叶片的基本生理过程得以维持。但高气孔密度在增加CO2吸收的同时,必然会增强水分蒸发,这容易造成植物“生理干旱”。小气孔开度除可减少水分流失外,还具有灵活的调节功能,有利于快速关闭,可在一定程度上减少水分散失,对高气孔密度可能造成的“生理干旱”进行补偿,这可能是气孔对强光环境作出的响应和适应[21]。可见,依赖于气孔开度和气孔密度的双重调节是植物对长期光照差异适应的一种重要体现。
4 结论本研究通过分析具有鳞形和刺形两种叶型的3种圆柏属植物,观测了短期光照变化(日变化)与长期光照差异(树冠阴面和阳面)下叶片的气孔特征及其相关性。结果表明,短期光照变化下,植物叶片的气孔开度主要依赖于保卫细胞宽度的调节;长期光照差异下,3种圆柏属植物气孔开度依赖于保卫细胞长度和保卫细胞宽度的双重调节。可见,气孔特征可以较好地反映植物对长期和短期光照适应性的差异。
| [1] |
HEDRICH R, SHABALA S. Stomata in a saline world[J]. Current Opinion in Plant Biology, 2018, 46: 87-95. DOI:10.1016/j.pbi.2018.07.015 |
| [2] |
DONG H, BAI L, ZHANG Y, ZHANG G, MAO Y, MIN L, XIANG F, QIAN D, ZHU X, SONG C P. Modulation of guard cell turgor and drought tolerance by a peroxisomal acetate-malate shunt[J]. Molecular Plant, 2018, 11(10): 1278-1291. DOI:10.1016/j.molp.2018.07.008 |
| [3] |
BRODRIBB T J, JORDAN G J. Water supply and demand remain balanced during leaf acclimation of Nothofagus cunninghamii trees[J]. New Phytologist, 2011, 192(2): 437-448. DOI:10.2307/41320497 |
| [4] |
HETHERINGTON A M, WOODWARD F I. The role of stomata in sensing and driving environmental change[J]. Nature, 2003, 424(6951): 901-908. DOI:10.1038/nature01843 |
| [5] |
姚静远, 李东升, 郭琳, 胡佳成. 植物叶表气孔日变化规律与叶厚参数的相关性初探[J]. 浙江农业学报, 2015, 27(1): 44-48. DOI:10.3969/j.issn.1004-1524.2015.01.09 YAO J Y, LI D S, GUO L, HU J C. Preliminary study on the correlation between plant stomata aperture and leaf thickness parameters[J]. Acta Agriculturae Zhejiangensis, 2015, 27(1): 44-48. DOI:10.3969/j.issn.1004-1524.2015.01.09 |
| [6] |
吴永华, 廖伟彪, 许宏刚, 张建旗, 李捷. 10种地被植物叶片气孔特征及其耐阴性研究[J]. 甘肃农业大学学报, 2011, 46(5): 95-100. DOI:10.3969/j.issn.1003-4315.2011.05.019 WU Y H, LIAO W B, XU H G, ZHANG J Q, LI J. Relationships between leaf stomatal characteristics of ten kinds of ground cover plants and their shade tolerance[J]. Journal of Gansu Agricultural University, 2011, 46(5): 95-100. DOI:10.3969/j.issn.1003-4315.2011.05.019 |
| [7] |
王荣, 郭志华. 不同光环境下枫香幼苗的叶片解剖结构[J]. 生态学杂志, 2007, 26(11): 1719-1724. DOI:10.3969/j.issn.1673-8225.2012.06.005 WANG R, GUO Z H. Responses of seedling leaf anatomical structure of Liquidambar formosana, a deciduous broadleaf tree, to different light regimes[J]. Chinese Journal of Ecology, 2007, 26(11): 1719-1724. DOI:10.3969/j.issn.1673-8225.2012.06.005 |
| [8] |
黎有有, 唐源江, 李菁, 周毅, 黄衡宇. 蛇足石杉的形态学及不同光照处理下气孔特征的研究[J]. 植物研究, 2009, 29(4): 411-416. LI Y Y, TANG Y J, LI J, ZHOU Y, HUANG H Y. Morphology of Huperzia serrata and its stomata characteristics under different light treatments[J]. Bulletin of Botanical Research, 2009, 29(4): 411-416. |
| [9] |
段贝贝, 赵成章, 徐婷, 郑慧玲, 冯威, 韩玲. 兰州北山不同坡向刺槐叶脉密度与气孔性状的关联性分析[J]. 植物生态学报, 2016, 40(12): 1289-1297. DOI:10.17521/cjpe.2016.0215 DUAN B B, ZHAO C Z, XU T, ZHENG H L, FENG W, HAN L. Correlation analysis between vein density and stomatal traits of Robinia pseudoacacia in different aspects of Beishan Mountain in Lanzhou[J]. Chinese Journal of Plant Ecology, 2016, 40(12): 1289-1297. DOI:10.17521/cjpe.2016.0215 |
| [10] |
达光文, 许宫堂, 周光军, 朱金玉. 祁连山东段林区主要树种蒸腾耗水特征研究[J]. 人民黄河, 2010, 32(9): 78-79. DOI:10.3969/j.issn.1000-1379.2010.09.034 DA G W, XU G T, ZHOU G J, ZHU J Y. Study on transpiration water consumption characteristics of main tree species in forest area in Eastern Qilian Mountains[J]. Yellow River, 2010, 32(9): 78-79. DOI:10.3969/j.issn.1000-1379.2010.09.034 |
| [11] |
朱玉怀, 张大才, 李双智. 西藏东达山3种嵩草属植物气孔特征沿海拔的变化[J]. 西北植物学报, 2017, 37(4): 105-113. ZHU Y H, ZHANG D C, LI S Z. Variations of stomatal characters for three species of Genus Kobresia along an elevational gradient in the Dongda Mountains of Tibet[J]. Acta Botanica Boreali-Occidentalia Sinica, 2017, 37(4): 105-113. |
| [12] |
邵邻相, 张均平, 刘艳. 8种柏科植物叶表皮的扫描电镜观察[J]. 浙江师范大学学报(自然科学版), 2008, 31(2): 195-200. DOI:10.3969/j.issn.1001-5051.2008.02.017 SHAO L X, ZHANG J P, LIU Y. Observation of leaves of eight Cupressaceae species by SEM[J]. Journal of Zhejiang Normal University(Natural Sciences), 2008, 31(2): 195-200. DOI:10.3969/j.issn.1001-5051.2008.02.017 |
| [13] |
吕洪飞. 紫竹梅、吊竹梅和鸭跖草气孔分布与比较[J]. 植物学通报, 2000, 17(4): 375-380. DOI:10.3969/j.issn.1674-3466.2000.04.013 LYU H F. Comparative study on the stomatic distribution on plants Setereasea purpurea Boom, Zebrina pendula Schnizl and Commelina communis Linn[J]. Chinese Bulletin of Botany, 2000, 17(4): 375-380. DOI:10.3969/j.issn.1674-3466.2000.04.013 |
| [14] |
SHIMAZAKI K I, DOI M, ASSMANN S M, KINOSHITA T. Light regulation of stomatal movement[J]. Annual Review of Plant Biology, 2007, 58(1): 219-247. DOI:10.1146/annurev.arplant.57.032905.105434 |
| [15] |
王萍, 邱念伟, 侯文雨, 宋晓旭, 舒凤月, 周峰. "光和K+对气孔开度的影响"实验设计再优化[J]. 植物生理学报, 2020, 56(4): 194-203. DOI:10.13592/j.cnki.ppj.2019.0516 WANG P, QIU N W, HOU W Y, SONG X X, SHU F Y, ZHOU F. Experimental design reoptimization for "The effects of light and K+ on stomatal opening"[J]. Plant Physiology Journal, 2020, 56(4): 194-203. DOI:10.13592/j.cnki.ppj.2019.0516 |
| [16] |
杨克彤, 常海龙, 陈国鹏, 俞筱押, 鲜骏仁. 兰州市主要绿化植物气孔性状特征[J]. 植物生态学报, 2021, 45(2): 187-196. DOI:10.17521/cjpe.2020.0257 YANG K T, CHANG H L, CHEN G P, YU X Y, XIAN J R. Stomatal traits of main greening plant species in Lanzhou[J]. Chinese Journal of Plant Ecology, 2021, 45(2): 187-196. DOI:10.17521/cjpe.2020.0257 |
| [17] |
王瑞丽, 于贵瑞, 何念鹏, 王秋凤, 赵宁, 徐志伟. 气孔特征与叶片功能性状之间关联性沿海拔梯度的变化规律——以长白山为例[J]. 生态学报, 2016, 36(8): 2175-2184. DOI:10.5846/stxb201411042162 WANG R L, YU G R, HE N P, WANG Q F, ZHAO N, XU Z W. Altitudinal variation in the covariation of stomatal traits with leaf functional traits in Changbai Mountain[J]. Acta Ecologica Sinica, 2016, 36(8): 2175-2184. DOI:10.5846/stxb201411042162 |
| [18] |
金鹰, 王传宽. 植物叶片水力与经济性状权衡关系的研究进展[J]. 植物生态学报, 2015, 39(10): 1021-1032. DOI:10.17521/cjpe.2015.0099 JIN Y, WANG C K. Trade-offs between plant leaf hydraulic and economic traits[J]. Chinese Journal of Plant Ecology, 2015, 39(10): 1021-1032. DOI:10.17521/cjpe.2015.0099 |
| [19] |
尹冰. 青藏高原常见植物叶片功能性状与气孔特征之间的关系及其变化规律[D]. 兰州: 兰州大学, 2018. YIN B. The relationship between leaf functional traits and stomatal characteristics of common plants in Tibetan Plateau[D]. Lanzhou: Lanzhou University, 2018. |
| [20] |
祁建, 马克明, 张育新. 辽东栎(Quercus liaotungensis)叶特性沿海拔梯度的变化及其环境解释[J]. 生态学报, 2007, 27(3): 930-937. DOI:10.3321/j.issn:1000-0933.2007.03.013 QI J, MA K M, ZHANG Y X. The altitudinal variation of leaf traits of Quercus liaotungensis and associated environmental explanations[J]. Acta Ecologica Sinica, 2007, 27(3): 930-937. DOI:10.3321/j.issn:1000-0933.2007.03.013 |
| [21] |
胡选萍, 吉成均, 安丽华. 青藏高原草地双子叶植物叶片的气孔特征研究[J]. 西北植物学报, 2015, 35(7): 1356-1366. DOI:10.7606/j.issn.1000-4025.2015.07.1356 HU X P, JI C J, AN L H. Leaf stomatic characteristics of dicotyledonous plants in the Tibetan Plateau grasslands[J]. Acta Botanica BorealiOccidentalia Sinica, 2015, 35(7): 1356-1366. DOI:10.7606/j.issn.1000-4025.2015.07.1356 |
(责任编辑 邹移光)