 2023, Vol. 50
2023, Vol. 50文章信息
引用本文 |

基金项目
- 安徽省科技重大专项(202103c08020006); 安徽省自然科学基金(2108085MC100); 安徽省农业科学院人才项目(QNYC-201903, QNYC-202103)
作者简介
- 江建华(1981—),男,博士,副研究员,研究方向为水稻杂种优势利用,E-mail:peanutlabjjh@163.com.
通讯作者
- 王德正(1962—),男,研究员,研究方向为水稻杂种优势利用,E-mail:wdzhengqin@163.com.
文章历史
- 收稿日期:2023-08-03

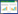


【研究意义】目前,我国杂交稻在稻米品质、病虫害抵御能力、氮高效利用和抗倒伏性等方面的研究一直处于瓶颈期,虽然经过育种家们多年协作攻关,选育出了一批占主导地位的两系新组合,但由于不育系来源均为安农S(正温敏核不育系的育性基因来源),其遗传背景单一、同质化问题严重,使得水稻产量增幅难以达到预期,且米质、抗性等重要农艺性状仍与社会需求存在差距。更为重要的是,两系核不育系育性易发生“飘移”,制种时母本遇低温导致其自交结实、制种失败[1-3]。加之当前全球气候异常变暖的大背景下,“冬暖夏凉”和“北暖南凉”时有发生,两系不育系“打摆子”的现象将常态化。寻求利用水稻杂种优势的育种新途径、新技术,乃是当今育种工作的关键所在。
【前人研究进展】杂交水稻育种方法可分为三系法、两系法和一系法。三系杂交水稻一般比普通良种增产20% 左右[4],两系杂交稻一般比同熟期的三系杂交水稻增产5%~10%[5]。目前,两系杂交水稻由于生产程序简化、杂交配组自由、可克服三系不育细胞质负效应和易实现籼粳亚种间杂种优势利用等特点而得到广泛应用。两系杂交稻品种在2007—2012年的推广面积位居全国首位[6]。随着Y两优系列、C两优系列、广占63系列、株两优系列杂交品种的推广,两系杂交稻品种在水稻生产中发挥着越来越重要的作用。
但水稻杂种优势利用存在的问题也日益凸显,如三系单产徘徊不前;缺乏强优势的早稻早、中熟组合;米质与抗性无突破性进展[7-8];杂交粳稻优势不突出;不育细胞质较单一;存在某种毁灭性病虫害暴发风险等问题。两系杂交水稻制种时其母本光温敏核不育系育性不稳定,光敏核不育现象受多个基因控制的遗传机理较为复杂,温度条件及遗传背景对育性转换也产生明显影响。其次是长江中下游地区要求安全的育性转换温度≤23.1 ℃。而一般籼稻繁殖临界致害温度为20 ℃。因此,适宜不育系种子繁殖的安全临界温度范围过窄[9]。再者,不育系的育性易发生“飘移”,增加了两系法制种的风险。
我国水稻三交种的育种始于20世纪70年代[10],是以水稻良种为临时保持系与纯三系不育系杂交,配制杂种不育系,然后将其再与恢复系杂交,获得杂种一代的一种培育杂交稻新途径。研究表明,部分三交种产量极显著高于对应的单交种,且认为三交种育种成败的关键在于亲本的正确选择和合理搭配[11]。三交种具有增强组合抗逆性、提高稳产性和制种产量以及加速扩大优良品种用于杂交稻育种等突出优点。但由于技术原因,三交种或多交种研究滞后,一定程度上限制了杂交水稻拓展遗传基础、增强适应性和提高稳产性,不利于进一步发掘杂交水稻的增产潜力。
目前,生产上大面积应用的杂交水稻均为温敏或光温互作类型,对反(光)温敏核不育系的研究较少。李训贞等[12]最早报道了反温敏核不育系N-10s和N-13s有关育性的鉴定结果,之后反温敏核不育系的相关报道主要集中在其本身对温、光反应特性的研究方面[13-14],未涉及反(光)温敏核不育系的育种利用研究。梁满中等[15]以反温敏核不育系go543S与光温敏核不育系农垦58S、7001S、培矮64S和长选3S配制杂交F1,结果表明,得到的F1在自然长日高温、短日低温和不同人工光、温处理条件下,花粉均表现不育、自交结实率为0,并未出现go543S或农垦58S类的育性转换。
近年来,随着科技进步,我国陆续开展了智能核不育系和永久温敏核不育系的研究,促进了两系法技术的进一步完善。但智能核不育系的应用由于涉及转基因技术和稻曲病为害而进展缓慢。
【本研究切入点】因用于临时保持系繁殖的不育系是杂种一代不育系与恢复系配组的三交F1,而用于配组的不育系实质上是上一交的F2,自然会产生某些分离[16],最终由于无法控制杂种株高、熟期的整齐性,使水稻三交杂种失去了应用价值。通过聚合水稻温敏和反温敏核不育基因,创制新型杂交水稻遗传工具——水稻永久温敏核不育系[17],不仅可解决现有温敏核不育系存在的育性问题[1-2],还可解决制种基地选择范围狭小和制种季节紧张等矛盾[18]。此外,还为广泛应用抗性、优质等有利基因培育多系杂交种带来方便,并且配组自由、不受恢保关系限制,有利于增强水稻抗逆性和适应性。【拟解决的关键问题】鉴于此,本研究在发现永久温敏核不育现象的基础上,构建了天丰B的温敏和反温敏核不育近等基因系,创制了水稻永久温敏核不育系,并对其育性遗传特性进行初步研究,旨在揭示永久温敏核不育遗传规律,以期为四系(温敏核不育系、反温敏核不育系、永久温敏核不育系和恢复系)杂交稻育种和研究提供理论依据。
1 材料与方法 1.1 试验材料水稻不育系26份:籼型温敏核不育系有4份,分别为矮占43S、1892S、广茉S和新二S;反温敏核不育系有1份,为雁农s;粳型光(温)敏核不育系有2份,分别为7001S和518S;反光(温)敏核不育系有1份,为粳5278s;天丰B温敏核不育近等基因系有7个,命名为天丰Zn材料,以Z1~Z7表示;天丰B反温敏核不育近等基因系有10个,命名为天丰Fn材料,以F1~F10表示;永久温敏核不育系1份,为天丰M。
恢复系材料有4份,分别为R998、R1392、R1501和R1032。
杂交种子有47份,分别是6个温敏核不育系和2个反温敏核不育系相互杂交收获的杂交种子12份(表 1);温敏(Thermo-sensitive genic male sterlie,TGMS)和反温敏核不育(Reverse thermo-sensitive genic male sterlie,RTGMS)近等基因系成对测交获得的测交种子31份,以C1~C31表示(表 2);天丰M(C8)与4个恢复系配组采集种子4份。
|
|
1.2 永久温敏核不育系天丰M的选育
参照王德正等[19]的方法,2010年春季在安徽省农业科学院水稻研究所海南南繁陵水基地(18°30′N,109°45′E),选用优质、多抗、综合性状优良的三系保持系天丰B(测交选育的天丰A配合力强)作为桥梁轮回亲本,分别与雁农s和矮占43S杂交。2010年夏季在安徽省农业科学院水稻研究所郭河试验基地(31°83′N,117°25′E)种植F1收获F2种子(图 1)。在适于温敏和反温敏核不育特性表达的环境压力下种植分离世代F2。从F2中选择主要性状偏向天丰B的不育单株,与天丰B回交,种植BC1F1收获种子。再从BC1F2中选择主要性状偏向天丰B的不育单株,与天丰B回交,并以此反复。通过回交和性状筛选,直至获得除不育特性外、其他性状与天丰B基本一致的温敏核不育近等基因系天丰Zn和反温敏核不育近等基因系天丰Fn,选育过程和效果见图 1、图 2。进而由天丰Zn姊妹系与天丰Fn姊妹系成对测交获得理想组合C8和C12,由此配制永久温敏核不育系天丰M(以母本第一拼音字母M为符号代替不育系,并与三系A和两系S区别之)。

|
| 图 1 利用天丰B作桥梁轮回亲本选育温敏和反温敏核不育近等基因系天丰Zn(Fn) 的过程 Fig. 1 Breeding process of a near-isogenic line Tianfeng Zn(Fn) of TGMS and RTGMS with Tianfeng B as a bridge recurrent parent |

|
| 图 2 天丰B温敏和反温敏核不育近等基因系与基因供体亲本株型比较 Fig. 2 Comparison of plant type between gene donor parents and near-isogenic lines of TGMS and RTGMS in Tianfeng B |
1.3 育性观测方法
1.3.1 不同温敏和反温敏核不育系杂交F1育性调查 2010年4月28日,将6个温敏核不育系和2个反温敏核不育系相互杂交的12个组合播种在安徽省农业科学院水稻研究所郭河试验基地试验田,30 d后移栽。自始穗期开始,每隔2~4 d各材料分别取5穗(已抽穗但尚未开花)套自交袋并标注日期,成熟后考查单穗总粒数和实粒数,计算自交结实率。8月10日从12个组合中分别取5株割茬移栽(留荐高度15~20 cm),每株剥蘖栽5穴,待抽穗未开花时每穴套自交袋,成熟后考查单穗总粒数和实粒数,计算自交结实率。计算公式如下:
自交结实率(%)=单穗实粒数/总粒数×100
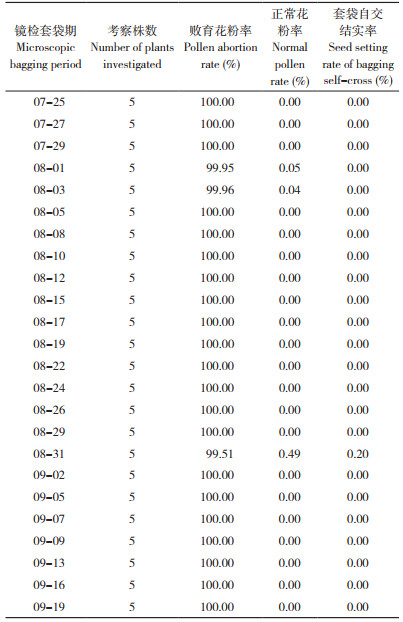
1.3.2 永久温敏核不育系天丰M的育性考查 2015年4月30日起,将永久温敏核不育系天丰M(C8)分8期播种于安徽省农业科学院水稻研究所郭河试验基地,每期隔10 d,播种25 d后移栽。自始穗期开始,每个播期每隔2~4 d取5穗,每穗取上、中、下3个部位已成熟但尚未开花的花药1个,用1% 的碘- 碘化钾溶液染色,镜检3个视野中的花粉染色情况,统计不同播期各材料的花粉败育率,直至抽穗结束;每次镜检时,各材料均选5个已抽穗但尚未开花的穗套自交袋并标注日期,成熟后考查单穗总粒数和实粒数,计算自交结实率。
1.3.3 永久温敏核不育系天丰M杂种优势测验 2016年春季在安徽省农业科学院水稻研究所海南南繁陵水基地利用永久温敏核不育系天丰M(C8),与R998等15个恢复材料杂交,手工配制杂交种15份,2016年夏季种植于安徽省农业科学院水稻研究所郭河试验基地,5月10日播种,6月10日单本移栽。各组合均种植3行,每行11株,株行距16.7 cm×26.7 cm。3次重复,随机区组排列,田间管理同一般大田。成熟时每小区从中间取5株于室内考查株高、有效穗数、穗长、每穗总粒数、每穗实粒数、每穗秕粒数(计算结实率),使用千分之一天平秤量千粒质量和单株谷质量。
1.3.4 正、反温敏核不育近等基因系测交F1育性观察 2017年夏季,在安徽省农业科学院水稻研究所郭河试验基地,以Z1~Z7为母本分别与F1~F10测交(Zn×Fn)获得测交组合31个,冬季12月10日在安徽省农业科学院水稻研究所海南南繁陵水基地播种,其中9个未出苗,其他于25 d后单苗移栽,每组合16~44株不等,株行距16.7 cm ×26.7 cm,厢宽133.3 cm,走道40 cm。抽穗后多次考察各组合单株表现,其中7个组合串粉混杂淘汰,最后统计数据组合15个。2018年3月12日对15组合中的不育株割茬让其再生,4月10日抽穗后观察低温不结实单株在高温下的自交结实情况。
2 结果与分析 2.1 水稻永久温敏核不育现象温敏和反温敏核不育性皆由隐性核不育基因控制,温敏核不育系与反温敏核不育系杂交的F1应像不等位的2个温敏核不育系杂交的F1一样,无论在长日高温还是在短日低温下都应表现可育。由表 1可知,籼型反温敏核不育系雁农s与籼型温敏核不育系广茉S、新二S杂交的F1,以及粳型反温敏核不育系粳5278s与粳型光(温)敏核不育系7001S、518S杂交的F1,自交结实率均达到38% 以上,完全符合2个不等位的隐性不育基因可相互恢复育性的规律。
此外,即便是温敏核不育基因来源相同、表现等位的2个温敏核不育系杂交的F1,如1892S×矮占43S、广茉S×新二S,虽然在长日高温下不育但在短日低温下还是表现可育,自交结实率高于15%(表 1),符合基因纯合材料育性随温度变化而变化的规律。
然而反温敏核不育系雁农s与2个温敏核不育系矮占43S、1892S杂交的F1(表 1),在高、低温条件下均自交不结实,表现为永久不育现象。但在粳型材料中却未发现这种现象,可以理解为目前应用的粳型温敏和反温敏核不育系中没有等位的温敏核不育基因。
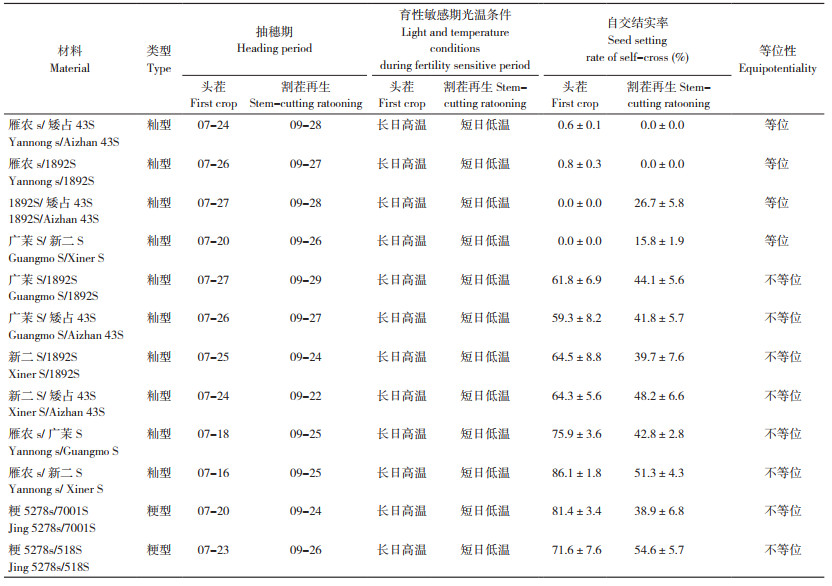
2.2 温敏和反温敏核不育近等基因系测交F1代育性分离规律在观察天丰M育性变化的试验中,发现不同测交组合的后代分离有差异,为明晰永久温敏核不育的遗传规律,揭示永久温敏核不育机制,参考三系不育系测配方法,2017年用温敏核不育系天丰Z姊妹系与反温敏核不育系天丰F姊妹系成对测交,分析F1代育性分离规律,15个温敏和反温敏核不育姊妹系测交组合(Zn ×Fn)F1代育性分离情况列入表 2。
从表 2可以看出,不同温敏和反温敏核不育姊妹系测交,在安徽省农业科学院水稻研究所海南南繁陵水基地低温下种植,F1出现可育、正常可育、永久不育和不育(植株类似永久不育,图中未展示)4种植株表型(图 3)。可育株株高与温敏和反温敏不育系相近,结实率与温敏和反温敏核不育系相当,且与育性敏感期温度密切相关(图 3B);正常可育株株高、结实率和繁茂性类似于品种间杂交种(图 3C);不育株株高与温敏和反温敏不育系相近,在高、低温下均不结实,结实率低于3%(图 3D);不育株株高与温敏和反温敏不育系相近,其育性随温度变化而变化,低温下不育、高温下可育。群体分离表现4种类型:

|
| A:天丰F;B:可育株;C:正常结实株;D:永久不育株;E:天丰Z A: Tianfeng F; B: Fertile plant; C: Normal seed-setting plant; D: Permanently sterile plant; E: Tianfeng Z 图 3 天丰Z与天丰F成对测交F1植株表型 Fig. 3 Plant phenotypes of F1 generation from cross between Tianfeng Z and Tianfeng F |
类型1,组合C8(Z6×F8)、C12(Z7×F10)在低温下所有植株不育,割茬再生仍保持不育,表现为永久不育,2个组合植株数分别为21株、18株。
类型2,C1(Z6×F1)、C3(Z6×F3)、C13(Z7×F5)、C16(Z7×F7)和C26(Z7×F9)5个组合在低温下表现可育与不育分离,不育株割茬再生仍保持不育,属永久不育株,各组合永久不育与可育植株数依次为9∶13、19∶25、13∶15、18∶22和10∶12,符合1对杂合基因1∶1的分离比例。
类型3,C4(Z1×F8)、C15(Z2×F8)、C17(Z2×F10)、C21(Z4×F8)、C24(Z5×F8)、C20(Z4×F10)、C28(Z1×F10)7个组合在低温下均表现不育,割茬再生发生不育与可育分离,各组合永久不育与低温不育高温可育株数依次为11∶9、16∶19、8∶8、12∶10、23∶19、11∶8和9∶7,也符合1对杂合基因1∶1的分离比例。
类型4,C31(Z1×F7)组合在低温下表现不育、可育株和正常可育,不育株割茬再生发生不育与可育分离,实际表型永久不育5株、低温不育高温可育6株、低温可育4株、正常可育7株,符合2对杂合基因1∶1∶1∶1分离比例。
从不同温敏和反温敏核不育系基因型预测F1的表现型列于表 3,表型预测与不同温敏和反温敏近等基因系测交F1代群体表现吻合。

|
据此分析测交4种表型(F1)及其父母本X位点的基因型如下:
类型1为XZXF,母本Z6、Z7为XZXZ,父本F8、F10为XFXF。
类型2为XZXF、XZXL,母本Z6、Z7为XZXZ,父本F1、F3、F5、F7、F9为XFXL。
类型3为XZXF、XFXL,母本Z1、Z2、Z4、Z5为XZXL,父本F8、F10为XFXF。
类型4为XZXF、XZXL、XFXL、XLXL,母本Z1为XZXL,父本F7为XFXL。
2.3 永久温敏核不育系天丰M的育性变化在天丰Zn与天丰Fn测交筛选组合而获得永久温敏核不育系天丰M的基础上,分期播种观察天丰M的育性变化,从始穗期(7月25日)至调查结束(9月19日),其花粉败育率均达到99.51% 以上,套袋自交结实率更是低于0.20%(表 4)。这表明天丰M无论在长日高温下还是在短日低温下均表现不育,称之为永久温敏核不育系。在由特定的温敏和反温敏核不育系杂交产生的永久核不育现象的基础上,通过桥梁亲本创制了永久温敏核不育系。
|
2.4 天丰M配组杂种优势
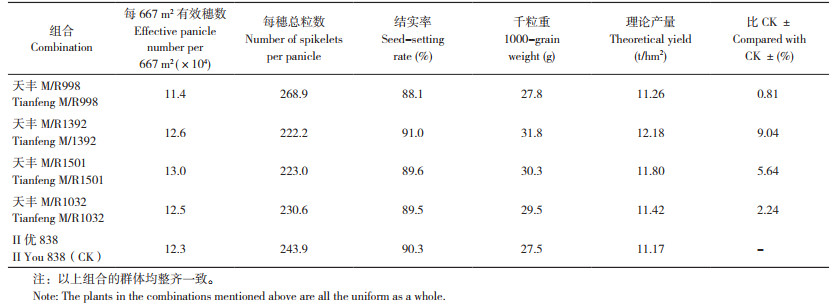
利用永久温敏核不育系天丰M与R998等15个恢复系材料测配,F1表现见表 5。永久温敏核不育系天丰M与恢复系R998、R1392、R1501和R1032杂交后代表现整齐一致,理论产量11.26~12.18 t/hm2,比对照种Ⅱ优838增产0.81%~9.04%,杂种优势明显。由此可见,当恢复系温敏核不育基因为显性时,与永久温敏核不育系天丰M杂交,不育性即可恢复,杂交F1表现可育、结实正常,不同组合表现不同的杂种优势,有些组合优势明显,推广应用前景可期。
3 讨论
本研究利用4个籼型温敏核不育系与1个籼型反温敏核不育系杂交以及2个粳型温敏核不育系与1个籼型反温敏核不育系杂交配制12个正反交F1组合。其中,籼型反温敏核不育系雁农s与籼型温敏核不育系广茉S、新二S杂交的F1,以及粳型反温敏核不育系粳5278s与粳型光(温)敏核不育系7001S、518S杂交的F1,自交结实率均达到38% 以上,表明上述(光)温敏核不育系中的不育基因与反温敏核不育系中的不育基因聚合后可以相互恢复育性。这些结果与课题组前期研究结果一致[17]。反温敏核不育系雁农s与2个温敏核不育系矮占43S、1892S杂交的F1,在高、低温条件下均自交不结实,表现为永久不育现象。但在粳型材料中却未出现这种现象,表明目前应用的粳型温敏和反温敏核不育系中没有等位的温敏核不育基因。
基于永久温敏核不育现象,通过选育桥梁轮回亲本天丰B温敏和反温敏核不育近等基因系,创制了永久温敏核不育系天丰M,并研究天丰M育性表现,确定其在高温或低温下均表现不育。这一发现尤其是不育种质创制,在开辟水稻杂种优势利用新途径的同时,规避了两系杂交水稻种子生产过程中因两用核不育系易受环境影响而带来的风险。此外,由于配组自由、不受恢保关系限制,其育性可被多数品种恢复,实用性与两系杂交稻等同。育性转化主导因子仍然是温度,但不同的是受温度调节的调控基因可在温敏和反温敏间切换,不育表现与现有的温敏核不育系包括温敏型(高温不育、低温可育)和反温敏型(低温不育、高温可育)研究结果一致[15, 20-23]。因此,永久温敏核不育系种子生产应视繁殖季节来确定父母本,如长江流域夏季繁殖,育性敏感期处于高温条件下,生产中应以温敏核不育近等基因系为母本,反温敏核不育近等基因系为父本。
随着新不育系的发现和新基因功能的研究,水稻光温敏雄性不育表现趋于复杂,其可能受多个基因调控,这些基因通过何种方式调控目的基因的表达、功能基因存在何种调控方式尚未有定论[6]。亚洲栽培稻种内存在极为丰富的功能等位基因自然变异,表现为超过1 040万个功能等位基因,平均每个基因具有226个功能等位基因[24]。本研究采用不同温敏和反温敏核不育姊妹系成对测交,分析了测交F1的育性分离规律。结果显示,只有1对基因控制永久温敏核不育,事实上控制温敏和反温敏核不育的结构基因在温敏和反温敏核不育系选育过程中,通过表型选择就已达纯合。因此,永久温敏核不育是由2对基因控制的,一对是温敏核不育基因,另一对是受温度调节的调控基因,起开关作用,主要调控不育结构基因。调控基因可能位于不同染色体上,也可能位于同一染色体的不同位点,从不同温敏和反温敏核不育姊妹系测交后代分离结果看,是位于某一染色体上的X位点、属于复等位基因,常态下,这个位点表现为对温度钝感或缺失,但来自温敏和反温敏核不育系的复等位基因互为显隐性关系,永久不育系在高温下,来自温敏核不育系的复等位基因为显性,调控结构基因表现不育;在低温下,来自反温敏核不育系的复等位基因切换为显性,同样调控结构基因表现不育,温敏和反温敏核不育系在高、低温下的育性转换也能证明这一点。
目前,永久温敏核不育系育性遗传特性研究虽取得了一些进展,但这些基因通过何种方式调控目的基因,遗传信息如何传导,以什么代谢物质调控不育性还不明确,仍需加强、深入研究。而调控基因为杂合状态,分别来源于温敏和反温敏核不育系,且互为显隐性关系,温度为主要调控因子等信息有助于了解其调控方式和温度受体。若温度受体就是热激蛋白(HSPs)[25],则有可能关联到高、低温危害水稻的研究。
4 结论针对温敏和反温敏核不育系杂交表现永久不育的现象,通过桥梁亲本聚合温敏和反温敏核不育基因育成水稻永久温敏核不育系,为杂种优势利用开辟了新途径、新方法。永久温敏核不育系育性稳定,在自然长日高温和短日低温下均表现不育,与温敏核不育系相比其优势就在于制种安全可靠。永久温敏核不育系配组自由,不像三系受恢保关系限制,有利于挖掘杂种优势潜力。永久不育特性缓解了制种季节和基地难选择的矛盾,且制种基地和扬花时段可以最大程度优化,为制种高产优质奠定了基础。此外,利用桥梁亲本、恢复系、温敏和反温敏核不育近等基因系,有利于水稻高产稳产。
| [1] |
CHEN L B, LEI D Y, TANG W B, XIAO Y H. Thoughts and practice on some problems about research and application of two-line hybrid rice[J]. Rice Science, 2011, 18(2): 79-85. DOI:10.1016/S1672-6308(11)60012-7 |
| [2] |
唐文帮, 陈良碧, 王建龙, 刘海, 肖应辉, 邓化冰, 刘国华. 水稻温敏型两用核不育系繁殖存在的问题及解决办法[J]. 杂交水稻, 2011, 26(5): 25-29. DOI:10.3969/j.issn.1005-3956.2011.05.008 TANG W B, CHEN L B, WANG J L, LIU H, XIAO Y H, DENG H B, LIU G H. Problems of and solutions to multiplication of TGMS line in rice[J]. Hybrid Rice, 2011, 26(5): 25-29. DOI:10.3969/j.issn.1005-3956.2011.05.008 |
| [3] |
王丰. 杂交水稻育种成就与展望——广东省农业科学院水稻研究所杂交水稻研究50年回顾[J]. 广东农业科学, 2020, 47(12): 1-11. DOI:10.16768/j.issn.1004-874X.2020.12.001 WANG F. Achievements and prospects of hybrid rice breeding-review of 50 year's research on hybrid rice by Rice Research Insitute of Guangdong Academy of Agricultural Sciences[J]. Guangdong Agricultural Sciences, 2020, 47(12): 1-11. DOI:10.16768/j.issn.1004-874X.2020.12.001 |
| [4] |
袁隆平. 杂交水稻的育种战略设想[J]. 杂交水稻, 1987(2): 1-3. DOI:10.16267/j.cnki.1005-3956.1987.01.001 YUAN L P. The tentative plan of hybrid rice breeding strategy[J]. Hybrid Rice, 1987(2): 1-3. DOI:10.16267/j.cnki.1005-3956.1987.01.001 |
| [5] |
袁隆平. 中国杂交水稻的研究与发展[J]. 科技导报, 2016, 34(20): 64-65. YUAN L P. Research and development of hybrid rice in China[J]. Science & Technology Review, 2016, 34(20): 64-65. |
| [6] |
黄惠芳, 鄂志国, 祁永斌, 管耀祖. 中国两系杂交稻的发展现状及光温敏雄性不育基因研究进展[J]. 浙江农业学报, 2015, 27(5): 893-899. DOI:10.3969/j.issn1004-1524.2015.05.30 HUANG H F, E Z G, QI Y B, GUAN Y Z. Current situation for development of two-line hybrid rice in China and research progress of rice photoperiod-and thermo-sensitive genic male sterility gene[J]. Acta Agriculturae Zhejianggensis, 2015, 27(5): 893-899. DOI:10.3969/j.issn1004-1524.2015.05.30 |
| [7] |
刘倩, 张国豪, 车万均, 肖瑶, 张杰, 胡运高. 杂交水稻重要亲本农艺性状配合力遗传力分析[J]. 广东农业科学, 2020, 47(1): 1-8. DOI:10.16768/j.issn.1004-874X.2020.01.001 LIU Q, ZHANG G H, CHE W J, XIAO Y, ZHANG J, HU Y G. Analysis on combining ability and heritability in agronomic traits of key parents of hybrid rice[J]. Guangdong Agricultural Sciences, 2020, 47(1): 1-8. DOI:10.16768/j.issn.1004-874X.2020.01.001 |
| [8] |
朱满山, 王丰, 符福鸿, 黄慧君, 肖昕, 柳武革, 刘振荣, 廖亦龙, 李金华, 陈建伟, 付崇允. 籼型杂交稻亲本米质性状的遗传效应对比分析[J]. 广东农业科学, 2012, 39(13): 1-5, 8. DOI:10.16768/j.issn.1004-874X.2012.13.001 ZHU M S, WANG F, FU F H, HUANG H J, XIAO X, LIU W G, LIU Z R, LIAO Y L, LI J H, CHEN J W, FU C Y. Comparative analysis of parental lines in genetic effect on grain quality traits in indica hybrid rice[J]. Guangdong Agricultural Sciences, 2012, 39(13): 1-5, 8. DOI:10.16768/j.issn.1004-874X.2012.13.001 |
| [9] |
雷东阳, 林勇, 陈立云. 水稻两用核不育系的研究现状与发展策略[J]. 湖南农业大学学报(自然科学版), 2019, 45(3): 225-230. DOI:10.13331/j.cnki.jhau.2019.03.001 LEI D Y, LI N Y, CHEN L Y. T h e r es e ar ch pr o gr ess a nd countermeasures of dual-purpose genic male sterile line[J]. Journal of Hunan Agricultural Univesity (Natural Sciences), 2019, 45(3): 225-230. DOI:10.13331/j.cnki.jhau.2019.03.001 |
| [10] |
杨仁崔. 对三交稻的一些初步认识[J]. 福建农业科技, 1982(2): 5. YANG R C. Some preliminary knowledge on three-way rice[J]. Fujian Agricultural Science & Technology, 1982(2): 5. |
| [11] |
苏孝道, 杨仁崔, 卢浩然. 杂交水稻三交种的优势表现及配合力[J]. 福建农学院学报(自然科学版), 1990, 19(1): 7. SU X D, YANG R C, LU H R. Heterosis and combining ability of three-hybrid rice[J]. Journal of Fujian Agriculture and Forestry University (Natural Science Edition), 1990, 19(1): 7-12. |
| [12] |
李训贞, 陈良碧, 周庭波. 新型低温不育水稻(N-10s, N-13s)育性的初步鉴定[J]. 湖南师范大学自然科学学报, 1991, 14(2): 376-378. LI X Z, CHEN L B, ZHOU T B. Preliminary observation of fertility changes in the new type low temperature sensitive male sterile rice N-10s and N-13s[J]. Journal of Natural Science of Hunan Normal University, 1991, 14(2): 376-378. |
| [13] |
吴厚雄, 李必湖, 向阳, 梁满中, 徐孟亮, 陈良碧. 低温敏核不育系水稻go543S育性对温、光的反应[J]. 生态学报, 2003, 23(3): 463-470. DOI:10.3321/j.issn:1000-0933.2003.03.009 WU H X, LI B H, XIANG Y, LIANG M Z, XU M L, CHEN L B. Effects of temperature and photoperiod on the fertility of low temperature sensitive genic male sterile rice[J]. Acta Ecology Sinica, 2003, 23(3): 463-470. DOI:10.3321/j.issn:1000-0933.2003.03.009 |
| [14] |
徐孟亮, 张俊, 龚曼, 梁满中, 陈良碧. 一个反向水稻两用核不育系育性对温度与光周期的反应[J]. 湖南师范大学自然科学学报, 2010, 33(4): 85-88. DOI:10.3969/j.issn.1000-2537.2010.04.017 XU M L, ZHANG J, GONG M, LIANG M Z, CHEN L B. Response of fertility of a new reverse P(T)GMS line to temperature and photoperiod in rice[J]. Journal of Natural Science of Hunan Normal University, 2010, 33(4): 85-88. DOI:10.3969/j.issn.1000-2537.2010.04.017 |
| [15] |
梁满中, 王晓辉, 吴厚雄, 陈良碧. 低温敏核不育系与光温敏核不育系杂交后代育性遗传的初步研究[J]. 作物学报, 2006, 32(10): 1537-1541. DOI:10.3321/j.issn.0496-3490.2006.10.018 LIANG M Z, WANG X H, WU H X, CHEN L B. Preliminary studies on heredity of fertility for low temperature induded genic male sterile line and photoperiod (thermo)-sensitive genic male sterile line in rice (Oryza sativa L.)[J]. Acta Agronomica Sinica, 2006, 32(10): 1537-1541. DOI:10.3321/j.issn.0496-3490.2006.10.018 |
| [16] |
李世英. 利用三交种杂种提高水稻竞争优势问题的争论[J]. 辽宁农业科学, 1986(2): 14-17. LI S Y. The debate on improving the competitive advantage of rice by using three-hybrid hybrid[J]. Liaoning Agricultural Sciences, 1986(2): 14-17. |
| [17] |
江建华, 倪金龙, 吴爽, 王德正. 聚合水稻温敏核不育基因和反温敏核不育基因创制永久核不育系[J]. 中国水稻科学, 2017, 31(4): 371-378. DOI:10.16819/j.1001-7216.2017.7020 JIANG J H, NI J L, WU S, WANG D Z. Develpment of permanet genic male sterile line by pyramiding thermo-sensitive genic male sterile genes and reverse temperature induced genic male sterile genes in rice (Oryza sativa L.)[J]. Chinese Journal of Rice Science, 2017, 31(4): 371-378. DOI:10.16819/j.1001-7216.2017.7020 |
| [18] |
肖层林, 张海清, 刘爱民, 佘雪晴. 两系法杂交水稻制种风险产生的原因及其控制对策[J]. 杂交水稻, 2010, 25(S1): 462-464. DOI:10.16267/j.cnki.1005-3956.2010.s1.053 XIAO C L, ZHANG H Q, LIU A M, SHE X Q. Causes of the risk in seed production of two-line hybrid rice and its controlling strategy[J]. Hybrid Rice, 2010, 25(S1): 462-464. DOI:10.16267/j.cnki.1005-3956.2010.s1.053 |
| [19] |
王德正, 吴爽, 杜士云, 王辉, 许学. 一种采用桥梁亲本选育水稻温敏永久核不育系的方法[P]. 中国专利: ZL201410065569.6, 2014-05-28. WANG D Z, WU S, DU S Y, WANG H, XU X. The invention relates to a method for breeding permanent thermo-sensitive genic male sterile lines in rice by using bridge parents[P]. China Patent: ZL201410065569.6, 2014-05-28. |
| [20] |
范优荣, 曹晓风, 张启发. 光温敏雄性不育水稻的研究进展[J]. 科学通报, 2016, 35(61): 3822-3832. DOI:10.1360/N972016-01047 FAN Y R, CAO X F, ZHANG Q F. Progress on photoperiod thermo-sensitive genic male sterile rice[J]. Chinese Science Bulletin, 2016, 35(61): 3822-3832. DOI:10.1360/N972016-01047 |
| [21] |
石明松. 对光照长度敏感的隐性雄性不育水稻的发现与初步研究[J]. 中国农业科学, 1985(2): 44-48. DOI:10.3864/j.issn.0578-1752.1985-18-02-44-48 SHI M S. The discovery and study of the photosensitive recessive male-ster ile rice (Oryza sativa L. subsp. Japonica)[J]. Scientia Agricultura Sinica, 1985(2): 44-48. DOI:10.3864/j.issn.0578-1752.1985-18-02-44-48 |
| [22] |
元生朝, 张自国, 许传桢. 光照诱导湖北光敏感核不育水稻育性转变的敏感期及其发育阶段的探讨[J]. 作物学报, 1988, 14(1): 7-13. YUAN S C, ZHANG Z G, XU C Z. Studies on the critical stage of fertility change induced by light and its phase development in HPGMR[J]. Acta Agronomica Sinica, 1988, 14(1): 7-13. |
| [23] |
刘宜柏, 贺浩华, 饶治祥, 孙义伟, 黄英金. 光温条件对水稻两用核不育系育性的作用机理研究[J]. 江西农业大学学报, 1991, 13(1): 1-7. DOI:10.13836/j.jjau.1991001 LIU Y B, HE H H, RAO Z X, SUN Y W, HUANG Y J. Studies of mechanism of fertility in dual-purpose genic male sterile line (O. sativa L.) under different light length and temperature conditions[J]. Acta Agriculturae Universitatis Jiangxiensis, 1991, 13(1): 1-7. DOI:10.13836/j.jjau.1991001 |
| [24] |
ZHANG F, WANG C C, LI M, CUI Y R, SHI Y Y, WU Z C, HU Z Q, WANG W S, XU J L, LI Z K. The landscape of gene-CDS-haplotype diversity in rice (Oryza sativa L.): Properties, population organization, footprints of domestication and breeding, and implications in genetic improvement[J]. Molecular Plant, 2021, 14(5): 787-804. DOI:10.1016/j.molp.2021.02.003 |
| [25] |
龙跃生, 陈良碧. 光敏、温敏雄性不育水稻基因研究进展[J]. 作物研究, 2001(2): 44-49. DOI:10.16848/j.cnki.issn.1001-5280.2001.02.019 LONG Y S, CHEN L B. Advances in studies of the genes for male sterility in photoperiod (thermo)-sensitive genic male sterile rice[J]. Crop Research, 2001(2): 44-49. DOI:10.16848/j.cnki.issn.1001-5280.2001.02.019 |
(责任编辑 马春敏)