 2023, Vol. 50
2023, Vol. 50文章信息
引用本文 |

基金项目
- 广东省自然科学基金(2021A1515010820);广东省现代农业产业技术体系专项(2023KJ105);广东省水稻育种新技术重点实验室项目(2023B1212060042)
作者简介
- 王晓飞(1982—),男,硕士,助理研究员,研究方向为水稻遗传育种,E-mail:wangxiaofei@gdaas.cn.
通讯作者
- 何秀英(1974—),女,博士,研究员,研究方向为水稻遗传育种,E-mail:xyhe@163.com.
文章历史
- 收稿日期:2023-11-03

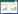


【研究意义】水稻是全球最重要的粮食作物之一,也是我国栽培面积最广、总产量最高的粮食作物。水稻生产对保障国家的粮食安全具有举足轻重的作用。然而,长期以来倒伏问题始终是水稻高产和优质化发展过程中的一大限制性因素。水稻如果发生大面积倒伏,将导致产量的严重损失和稻米品质的大幅下降,同时也会显著增加收割成本[1-2]。全球每年因作物倒伏所造成的损失巨大[3]。超级稻育种通常以大幅度提高生物量及株高来实现高产,大大增加水稻倒伏的风险。因此,提高水稻的抗倒伏性对水稻超高产育种至关重要[4]。当前,在水稻生产向机械化、轻简化、规模化发展的大趋势下,对水稻品种的抗倒伏性也提出了更高的要求[5]。因此,开展水稻的抗倒伏研究具有十分重要的理论意义和现实意义。【前人研究进展】为提高水稻抗倒伏性,必须确定抗倒伏相关性状,并建立客观、科学的评价方法[6]。目前,普遍采用通过测量基部茎秆抗折力的茎秆力学评价法来快速评估水稻品种的抗倒伏潜力[7-8]。大量研究表明,水稻基部茎秆抗折力与茎秆物理化学性状密切相关[9-10]。水稻茎秆由节和节间组成,其中节可以增强茎秆刚度,提高其抵抗外部弯曲力矩的能力。节间则由表皮、机械组织、薄壁组织和维管束等组成。茎秆的机械强度主要取决于其粗细和节间长短,同时还受基部茎秆组织解剖结构(维管束、机械组织等)及钾、硅等化学物质的影响[10-12]。研究表明,茎秆充实度越高、茎秆越粗、节间越短、茎秆壁越厚,则茎秆的机械强度就越大,抗折力就越强,同等条件下抗倒伏能力也就越强[13-15]。水稻基部茎秆抗折力还受其他性状的影响,如分蘖夹角、穗型等。此外,栽培方式等农业措施也会通过改变植株的形态结构特征影响水稻的抗倒伏性[15-18]。株高的增加会使植株重心提高,从而增加基部茎秆受力力矩和力臂的长度。当受力力矩大于植株的抗折力矩时,就会发生倒伏。增加水稻抗倒伏能力的关键在于增加基部茎秆抗折力,可从增加基部茎秆壁厚等方面考虑,从而达到高产且抗倒伏的要求[19]。【本研究切入点】水稻基部茎秆抗折力是水稻抗倒伏性的核心指标之一[20],但其与基部茎秆相关物理性状之间的关系尚不明确,抗倒伏遗传机制也尚不清楚。粤禾丝苗(YHSM)是广东省农业科学院水稻研究所抗病育种研究室近年来精心选育的优良常规水稻新品种,为当前广东省及广州市主导品种,该品种高产、优质、抗病,具有极强的抗倒伏性,多年来在多地种植均未发生倒伏。作为抗倒伏育种亲本,粤禾丝苗具有综合性状优良、配合力高、恢复性好等特点,已通过广东(2014)、江西(2017)、湖北(2019)、安徽(2021)等省品种审定并获得大面积生产应用。因此,本研究以粤禾丝苗为材料,探究基部茎秆主要物理性状与茎秆抗折力的相关性,找出与基部茎秆抗折力最相关的核心关键性状。【拟解决的关键问题】研究水稻基部茎秆抗折力与相关物理性状之间的关系,阐明粤禾丝苗的抗倒伏遗传机制。本研究对粤禾丝苗及其遗传分离群体的抗倒伏性及相关性状进行了测定和统计分析,为揭示其抗倒伏遗传机制和调控机制奠定基础。
1 材料与方法 1.1 遗传分离群体构建以自主育成的抗倒伏品种粤禾丝苗和广东省生产中广泛应用的易倒伏品种象牙香占(XYXZ)为亲本,构建遗传群体。2018年4月于广东省农业科学院水稻研究所大丰基地种植粤禾丝苗和象牙香占,每个重复各1个小区,每小区5行、每行20株,株行距均为18 cm,设置3个重复(图 1)。以粤禾丝苗为母本、象牙香占为父本进行杂交,获得F1代种子。2018年8月种植F1代20株(株行距16.67 cm×16.67 cm),成熟后测量抗倒伏相关性状,之后自交收获F2代种子。2019年4月于广东省农业科学院清远三坑水稻实验基地种植F2群体,共900株,双行种植(行间距为16.67 cm、33.33 cm交替,如图 2)。抽穗后20 d左右,随机选取生长正常的350株材料测量抗倒伏相关性状,然后采取单粒传的方法构建重组自交系,目前已获得稳定的重组自交系群体F8。

|
| 图 1 亲本材料粤禾丝苗(左)和象牙香占(右)的田间种植情况 Fig. 1 Field planting conditions of parent materials Yuehesimiao (left) and Xiangyaxiangzhan (right) |

|
| 图 2 F2代材料种植情况 Fig. 2 Planting conditions of F2 test materials |
1.2 抗倒伏力学数据测定
2018年6月对2个亲本及F2代群体抗倒伏相关性状进行测量。在齐穗后20 d早上8:00—10:00,对2个亲本及F2代群体主茎进行取样,每小区随机选择10株生长正常的植株,尽量保持不失水,测定以下抗倒伏相关指标:单株抗弯力(Brip)、主茎鲜重(MFw)、株高(Ph)、N2抗折力(Brs)、N3抗折力、N2长度(L)、N3长度、N2茎粗(Sd)、N3茎粗、N2长轴(Ld)、N3长轴、N2短轴(Sa)、N3短轴、N2壁厚(Wt)、N3壁厚、N2鲜重(Fw)、N3鲜重、N2充实度(Fd)、N3充实度。其中,N2、N3分别表示水稻基部茎秆第二、第三节间,其长、短轴分别表示节间类椭圆横切面的长、短轴。
1.2.1 单株抗弯力及株高测定 于植株距地面15 cm处,用YYD-1A茎秆强度测量仪垂直于茎秆向前推压,将稻茎压弯至45°倾角处(用量角器测),记录最大压力值,即为水稻单株最大折力,亲本材料每小区测定10株,取平均值为表型值。用1 500 mm钢尺测量植株从地面至穗尖的高度,取10株平均值为株高。
1.2.2 茎秆抗折力及茎粗测定 参考濑古秀生[21]方法,自行设计测定抗折力的简单支架,将待测定的节间置于支点间距离5 cm的测定器上(短于5 cm不测),该节间中点与测定器中点对应,用数字植株倒伏测定仪(YYD-1A)在节间中部匀速用力下压,直至茎秆折断,计数仪器数值即为该节间茎秆的抗折力,取10株平均值。
随机取10株主茎剥去叶鞘,用电子游标卡尺测定茎基5 cm茎秆最宽处宽度和最细处宽度,取平均值为茎粗。即茎粗(mm)=(长轴长+ 短轴长)/2。
1.2.3 N2长度和茎秆充实度 用直尺测量N2长度。用解剖刀及电子游标卡尺测定N2类椭圆横切面长轴、短轴的长度及壁厚,并用烘干法测定N2干重,计算茎秆充实度(单位长度茎秆干重)。假定水稻茎秆为圆杆型结构,为方便计算,充实度可以简单的以单位长度重量来体现。N2单位长度干重(mg/cm)=N2干重/N2长度。茎秆充实度=m/L(m代表所测量茎秆的鲜重,L表所测茎秆长度)
1.2.4 茎秆横切面组织染色及显微结构观察 采用间苯三酚染色法测定。将4 g间苯三酚溶于100 mL 95%乙醇中制备成染色液,将各材料徒手切片后选取完整薄片置于载玻片上,滴加染色液和浓盐酸各1滴,盖上盖玻片在显微镜下观察拍照,可见木质化细胞壁被染成红色。在显微镜下观察间苯三酚染色过的切片,观察机械组织细胞壁加厚情况。根据显微拍照结果测量茎秆壁厚,比较各品种水稻茎秆细胞结构的差异。
1.3 数据处理与分析试验数据采用IBM SPSS Statistics 25.0软件进行统计分析及部分图表绘制,部分图表利用Excel 2019软件进行绘制。
2 结果与分析 2.1 亲本及F2群体抗倒伏相关测量数据F2群体N2、N3抗倒伏相关测量数据如表 1所示,粤禾丝苗的大部分抗倒伏相关指标表现明显优于象牙香占和F2群体。在茎粗、抗折力、主茎鲜重及壁厚等方面,粤禾丝苗较象牙香占均具有一定的优势。表明粤禾丝苗可能具有更强的抗倒伏能力。F2群体的抗倒伏相关指标数值位于粤禾丝苗与象牙香占两个亲本之间。据此推断,抗倒伏性表现为粤禾丝苗 > F2群体 > 象牙香占。
|
由表 1可知,粤禾丝苗的N2茎粗(6.21±0.19 mm)、N3茎粗(5.75±0.18 mm)显著大于象牙香占的N2茎粗(4.60±0.09 mm)、N3茎粗(4.30±0.08 mm),表明粤禾丝苗的茎秆更为粗壮,有助于提高抗倒伏能力。粤禾丝苗的单株抗弯力(19.86±3.52N)显著高于象牙香占的单株抗弯力(5.01±1.07 N)。粤禾丝苗的主茎鲜重(22.07±1.00 g)显著高于象牙香占主茎鲜重(14.68±0.99 g)。象牙香占的株高(121.60±1.13 cm)略高于粤禾丝苗的株高(107.70±0.90 cm),但该因素对抗倒伏能力的影响需要进一步研究。粤禾丝苗的N2抗折力(16.58±0.48 N)、N3抗折力(12.71±0.70 N)均显著高于象牙香占的N2抗折力(9.93±0.72 N)、N3抗折力(5.22±0.91 N),表明粤禾丝苗基部茎秆具有更好的抗折性。粤禾丝苗的N2充实度(0.19±0.01 g/cm)、N3充实度(0.13±0.01 g/cm)均显著高于象牙香占的N2充实度(0.11±0.00 g/cm)、N3充实度(0.08±0.00 g/cm),表明粤禾丝苗的茎秆组织更为紧实,有助于提高抗倒伏能力。综合以上数据分析,我们认为粤禾丝苗的抗倒伏性优于象牙香占,具体表现在粤禾丝苗在茎粗、单株抗弯力、主茎鲜重、N2和N3的抗折力以及充实度等方面均显示出较大优势。
2.2 茎秆组织结构对粤禾丝苗抗倒伏性的影响众多研究表明,基部节间的物理结构是水稻抗倒伏的物质基础[22-23]。为揭示粤禾丝苗抗倒伏的物质基础,我们对粤禾丝苗和象牙香占基部N2节间进行细胞学分析,结果(图 3)显示,粤禾丝苗基部节间壁更厚,主要表现为薄壁细胞数目多,且机械组织和维管束鞘细胞间苯三酚染色着色深、木质化程度高;象牙香占薄壁组织少、机械组织和维管束鞘细胞间苯三酚染色着色浅,表明粤禾丝苗机械组织的纤维素含量高于象牙香占。

|
| 图 3 粤禾丝苗(A)和象牙香占(B)基部茎秆壁显微切片 Fig. 3 Micro-sections of the basal stem wall of Yuehesimiao (A) and Xiangyaxiangzhan (B) |
2.3 粤禾丝苗抗倒伏性的遗传分析
从900株粤禾丝苗×象牙香占的F2分离群体中随机选取350株进行抗倒伏相关性状的测量和统计分析,结果(图 4)发现,在F2群体中,N2抗折力(图 4A)、N2长度(图 4B)、N2茎粗(图 4C)、茎秆壁厚(图 4D)、鲜重(图 4E)和充实度(图 4F)均呈连续分布,除鲜重和充实度外均集中在双亲的平均水平、基本呈正态分布。因此,可以判断上述抗倒伏相关性状符合数量性状分布特征,由多个数量性状基因控制。

|
| 图 4 F2群体抗倒伏相关性状频率分布(n=350) Fig. 4 Frequency distribution of lodging resistance related traits of the F2 population (n=350) |
对N2抗折力、长度、茎粗、壁厚、鲜重、充实度等指标进行频率统计分析发现,N2抗折力(图 4A)在群体中的频率分布呈双峰形态,表明该群体中存在抗性较强和抗性较弱两个类型,为进一步明确抗性基因位点及其显隐性关系,需要开展深入研究。N2长度的频率分布峰值出现在8.4 cm附近,这个数值位于两个亲本之间(图 4B);大多数样本的N2长度数值分布在峰值左侧,表明F2群体N2长度表现更倾向于粤禾丝苗。N2茎粗的频率分布呈现单峰形态,峰值位于5.9~6.1 mm,基本上呈正态分布(图 4C)。N2壁厚在群体中的频率分布同样呈单峰形态,峰值位于0.70 mm附近;F2群体中茎秆壁厚相对均匀,符合正态分布特征。N2鲜重频率峰值在0.84 g,基本呈正态分布(图 4E)。N2充实度频率分布峰值集中在0.10~0.13 g/cm附近(图 4F)。
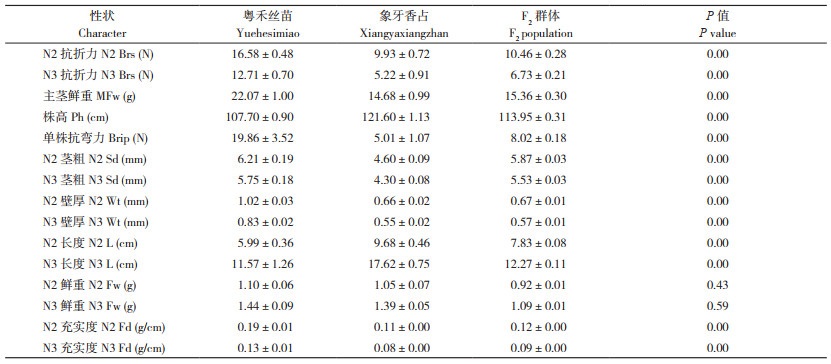
2.4 抗倒伏性状间的相关性分析本研究对粤禾丝苗×象牙香占的F2群体的N2抗折力、长度、茎粗、壁厚、鲜重、充实度等13个抗倒伏性状进行Pearson相关性分析,结果见表 2。由表 2可知,N2抗折力与N2壁厚(r=0.75**)、N2充实度(r=0.67*)、N2短轴(r=0.52)、主茎鲜重(r=0.50)、N2鲜重(r=0.48)相关性较显著;与N2茎粗(r=0.38)、株高(r=0.18)、N2长轴(r=0.17)相关性不显著,表明节间壁厚、充实度、短轴、茎秆粗细、节间长度均为N2抗折力的主要影响因素,应进一步深入研究。
|
我们还对N2与N3抗倒伏主要性状进行Pearson相关性分析,结果(表 3)表明,N2抗折力与N3抗折力(r=0.76**)、N2茎粗与N3茎粗(r=0.80**)、N2长度与N3长度(r=0.66**)、N2壁厚与N3壁厚(r=0.74**)、N2鲜重与N3鲜重(r=0.70**)、N2充实度与N3充实度(r=0.82**),N2、N3的多个相同性状呈现极显著正相关。而N2长度与N2充实度(r=-0.39*)、N3充实度(r=-0.25)呈负相关,N3长度与N2充实度(r=-0.47*)、N3充实度(r=-0.40*)呈显著负相关,表明N2和N3抗倒伏性状具有一定的遗传连锁关系。

|
3 讨论 3.1 水稻茎秆抗倒伏机制分析
本研究结果表明,N2壁厚与N2抗折力呈极显著正相关关系(r=0.75**)。N2壁厚越大,茎秆抗折力越强,抗倒伏性也越强,是水稻抗倒伏的核心性状。N2充实度与N2抗折力呈显著正相关(r=0.67*)。N2短轴与N2抗折力也呈正相关(r=0.52)。N2鲜重与N2抗折力呈正相关(r=0.48)。N2茎粗与N2抗折力呈正相关(r=0.38)。株高与N2抗折力呈弱正相关(r=0.18)。N2长轴与N2抗折力呈正相关(r=0.17);N2长度与N2抗折力呈负相关(r=-0.19)。N2长度与N2茎粗呈负相关(r=-0.20)。在各抗倒伏性状的相关性方面,N3与N2情况类似。
水稻基部茎秆节间壁厚、茎粗、充实度、长度等性状对抗倒伏性具有显著影响[24]。研究粤禾丝苗与象牙香占茎秆壁厚、充实度等性状之间的差异以及这些性状与抗折力的关系,有助于深入了解品种间抗倒伏性差异的机制[25-26]。由表 1可知,粤禾丝苗N2和N3的壁厚、充实度、茎粗以及主茎鲜重等抗倒伏性状指标均显著大于象牙香占,而N2长度、N3长度等指标则显著小于象牙香占。在抗倒伏直观性状方面,单株抗弯力表现为粤禾丝苗(19.86±3.52 N) > 象牙香占(5.01±1.07 N),N2抗折力表现为粤禾丝苗(16.58±0.48 N) > 象牙香占(9.93±0.72 N),这与表 2、表 3相关系数分析结果完全相符,即抗倒伏性较强的水稻品种通常具有较大的基部茎秆壁厚、较短的节间长度和较大的茎粗[27-30]。
由表 1可知,F2群体各项抗倒伏性状数值介于两个亲本之间,并未表现出偏向某一个亲本。图 4显示,F2群体的N2抗折力、长度、茎粗、壁厚、鲜重、充实度等性状频率分布基部符合正态分布,因此可以推断这些抗倒伏主要性状均为数量性状,由多基因控制。这一重要结论,可为今后抗倒伏遗传研究、分子机制研究奠定重要基础并明确研究方向,同时也可为水稻抗倒伏育种提供重要的理论依据。
3.2 水稻抗倒伏研究新思路水稻抗倒伏性是一项复杂的数量性状,受多个形态结构和生理生化因素的共同影响[31]。本研究发现,N2壁厚与N2抗折力呈极显著的正相关(r=0.75**),是决定茎秆抗折力的核心性状。因此,将研究重点聚焦在N2壁厚这个核心性状上,有效降低了水稻抗倒伏研究的复杂性,同时提高了研究目的性与效率,可为后续育种表型选择及基因功能分析奠定扎实的研究基础。对N2壁厚的深入研究,为揭示水稻抗倒伏分子机制、开展抗倒伏分子育种提供了可行性。
本研究中,株高与N2抗折力呈弱正相关(r=0.18),表明在一定范围内,随着株高的增加,水稻的抗折力也可能随之增大,这一研究结果对水稻矮化育种及抗倒伏育种理论是一次重要的补充和完善,具有重要意义。在现有基础上大幅提高水稻产量,必须大幅提高生物产量以获得更多的经济产量,这意味着增加株高是不可避免的。因此,超高产育种的主要途径应以提高茎秆抗折力为前提,在避免倒伏的情况下适当增加株高,从而提高生物产量和经济产量[32-33],这一策略为水稻超高产育种研究指明了方向。
3.3 水稻抗倒伏育种应用本研究结果可直接应用于水稻抗倒伏育种研究,在实际育种工作中具有很强的可操作性。选择具有以下特征的材料作为育种亲本有助于培育出综合性状优良的抗倒伏水稻新品种:(1)茎秆壁较厚:茎秆壁厚的植株具有较强的抗折断能力,可以抵御风力和重力的影响[34-37]。(2)基部茎秆节间较短:较短的节间可降低茎秆所受的弯折力矩,提高抗折力。(3)直径较粗:直径较粗的茎秆具有更高的机械强度,能够承受风雨等自然灾害冲击。(4)充实度高:茎秆内部的组织发育程度高,维管束较多、纤维素与木质素含量较高能增强茎秆的机械强度和耐受性[38-39]。
4 结论粤禾丝苗×象牙香占的F2群体抗倒伏性及其遗传机制研究表明:(1)水稻茎秆抗折力与N2基部节间茎秆壁厚呈极显著正相关,是茎秆抗倒伏的核心性状。(2)N2壁厚是一个数量性状,由多基因控制。N2壁厚越大则抗折力越强;N2抗折力与N2充实度、N2茎粗等性状呈正相关,而与N2长度呈负相关。(3)以N2壁厚为研究重点,可以大大降低以往水稻抗倒伏研究方法的复杂程度,大幅提高水稻抗倒伏研究的效率。
| [1] |
LUO X Y, WU Z F, LU F, DAN Z W, YUAN Z Q, LIANG T, ZHU R S, HU Z L, WU Z T. Evaluation of lodging resistance in rice based on an optimized parameter from lodging index[J]. Crop Science, 2022, 62(3): 1318-1332. DOI:10.1002/csc2.20712 |
| [2] |
刘佳欣, 吴周周, 周婵婵, 阿娜, 李漪濛, 王术. 水稻倒伏性状与抗倒途径研究进展[J]. 中国稻米, 2023, 29(6): 44-48, 55. DOI:10.3969/j.issn.1006-8082.2023.06.009 LIU J X, WU Z Z, ZHOU C C, A N, LI Y M, WANG S. Research progress of lodging characters and lodging resistance pathways in rice[J]. China Rice, 2023, 29(6): 44-48, 55. DOI:10.3969/j.issn.1006-8082.2023.06.009 |
| [3] |
齐龙昌, 周桂香. 水稻抗倒伏性状影响因素研究进展[J]. 安徽农业科学, 2019, 47(9): 19-22, 25. DOI:10.3969/j.issn.0517-6611.2019.09.006 QI L C, ZHOU G X. Advances in rsearch on fctors fnfluencing lodging resistance in rice[J]. Journal of Anhui Agricultural Sciences, 2019, 47(9): 19-22, 25. DOI:10.3969/j.issn.0517-6611.2019.09.006 |
| [4] |
滕祥勇, 王金明, 李鹏志, 林秀云, 孙强. 水稻抗倒伏性的影响因素及评价方法研究进展[J]. 福建农业学报, 2021, 36(10): 1245-1254. DOI:10.19303/j.issn.1008-0384.2021.10.018 TENG X Y, WANG J M, LI P Z, LIN X Y, SUN Q. Advances on studies relating to lodging resistance of rice plant[J]. Fujian Journal of Agricultural Sciences, 2021, 36(10): 1245-1254. DOI:10.19303/j.issn.1008-0384.2021.10.018 |
| [5] |
LIU Q H, MA J Q, ZHAO Q L, ZHOU X B. Physical traits related to rice lodging resistance under different simplified-cultivation methods[J]. Agronomy Journal, 2018, 110(1): 127-132. DOI:10.2134/agronj2017.09.0548 |
| [6] |
王晓飞, 刘斌. 水稻抗倒伏性遗传研究现状[J]. 广东农业科学, 2010, 37(7): 5-8. DOI:10.16768/j.issn.1004-874X.2010.07.039 WANG X F, LIU B. Progress on the genetics of lodging resistance in rice (Oryza sativa L.)[J]. Guangdong Agricultural Sciences, 2010, 37(7): 5-8. DOI:10.16768/j.issn.1004-874X.2010.07.039 |
| [7] |
LI Z Z, DENG F, ZHANG C, ZHU L, HE L H, ZHOU T, LU H, ZHU S L, ZENG Y L, ZHONG X Y, ZHOU W, CHEN Y. Can 'relative culm wall thickness' be used to evaluate the lodging resistance of rice?[J]. Archives of Agronomy and Soil Science, 2023, 69(6): 934-947. DOI:10.1080/03650340.2022.2046266 |
| [8] |
LIANG L, ZHANG X J, LIANG J, GUO Y M. Fuzzy synthetic evaluation on stalk-like crops with biomechanical properties indices[J]. Forest Chemicals Review, 2021, 330-340. DOI:10.17762/jfcr.vi.133 |
| [9] |
SHAH L, YAHYA M, SHAH S M A, NADEEM M, ALI A, ALI A, WANG J, RIAZ M W, REHMAN S, WU W, KHAN R M, ABBAS A, RIAZ A, ANIS G B, SI H, JIANG H, MA C. Improving lodging resistance: Using wheat and r ice as classical examples[J]. International Journal of Molecular Sciences, 2019, 20(17): 4211. DOI:10.3390/ijms20174211 |
| [10] |
ZHAO X Y, ZHOU N, LAI S K, FREI M, WANG Y X, YANG L X. Elevated CO2 improves lodging resistance of rice by changing physiochemical properties of the basal internodes[J]. Science of the Total Environment, 2018, 647: 223-231. DOI:10.1016/j.scitotenv.2018.07.431 |
| [11] |
JINGER D, DEVI M T, DHAR S, DASS A, SHARMA V K, VIJAYA KUMAR S, JOSHI E, JATAV H S, SINGH N. Silicon application mitigates abiotic stresses in rice: A review[J]. The Indian Journal of Agricultural Sciences, 2020, 90: 2043-2050. DOI:10.56093/ijas.v90i11.108557 |
| [12] |
YIN X H, TAO Z, HUANG M, ZOU Y B. Increasing wall thickness is more effective than increasing diameter for improving breaking resistance of rice internode[J]. Journal of Plant Biology and Crop Research, 2018(1001): 1-4. DOI:10.33582/2637-7721/1001 |
| [13] |
ASANO K, YAMASAKI M, TAKUNO S, MATSUOKA M. Artificial selection for a green revolution gene during japonica rice domestication[J]. Proceedings of the National Academy of Sciences of the United States of America, 2011, 108(27): 11034-11039. DOI:10.1073/pnas.1019490108 |
| [14] |
ARINICHEVA I V, DARMILOVA Z D. Construction of a mathematical model of cereal lodging // IOP Conference Series: Earth and Environmental Science[C]. IOP Publishing, 2021, 699(1): 012046. DOI: 10.1088/1755-1315/699/1/012046.
|
| [15] |
KASHIWAGI T, ISHIMARU K. Identification and functional analysis of a locus for improvement of lodging resistance in rice[J]. Plant Physiology, 2004, 134(2): 676-683. DOI:10.1104/pp.103.029355 |
| [16] |
OIKAWA T, KOSHIOKA M, KOJIMA K, YOSHIDA H, KAWATA M. A role of osga20ox1, encoding an isoform of gibberellin 20-oxidase, for regulation of plant stature in rice[J]. Plant Molecular Biology, 2004, 55(5): 687-700. DOI:10.1007/s11103-004-1692-y |
| [17] |
徐正进, 张树林, 周淑清, 刘丽霞. 水稻穗型与抗倒伏性关系的初步分析[J]. 植物生理学通讯, 2004, 40(5): 561-563. DOI:10.13592/j.cnki.ppj.2004.05.008 XU Z J, ZHANG S L, ZHOU S Q, LIU L X. Primary analysis of relationship between rice panicle type and lodging resistance[J]. Plant Physiology Communications, 2004, 40(5): 561-563. DOI:10.13592/j.cnki.ppj.2004.05.008 |
| [18] |
潘典进, 侯玲, 罗冬玉, 吴向阳, 郑文楼, 龚元春, 王军民. 籼型杂交水稻几个抗倒伏性状表现[J]. 湖北农业科学, 2019, 58(1): 15-18. DOI:10.14088/j.cnki.issn0439-8114.2019.01.003 PAN D J, HOU L, LUO D Y, WU X Y, ZHENG W L, GONG Y C, WANG J M. Several character expressions of lodging resistance of indica hybrid rice[J]. Hubei Agricultural Sciences, 2019, 58(1): 15-18. DOI:10.14088/j.cnki.issn0439-8114.2019.01.003 |
| [19] |
孙羽, 曾宪楠, 王麒, 宋秋来, 王曼力, 李希臣, 冯延江. 水稻倒伏影响因素及解决策略研究进展[J]. 黑龙江农业科学, 2023(9): 132-137. DOI:10.11942/j.issn1002–2767.2023.09.0132 SUN Y, ZENG X N, WANG Q, SONG Q L, WANG M L, LI X C, FENG Y J. Research progress on influencing factors and solving strategies of lodging resistance in rice[J]. Heilongjiang Agricultural Sciences, 2023(9): 132-137. DOI:10.11942/j.issn1002–2767.2023.09.0132 |
| [20] |
庞战士. 水稻茎秆性状与抗倒伏的相关性研究[J]. 园艺与种苗, 2022, 42(3): 58-59, 65. DOI:10.16530/j.cnki.cn21-1574/s.2022.03.025 PANG Z S. Correlation between stem traits and lodging resistance in rice[J]. Horticulture & Seed, 2022, 42(3): 58-59, 65. DOI:10.16530/j.cnki.cn21-1574/s.2022.03.025 |
| [21] |
濑古秀生. 水稻の倒伏に関する研究[J]. 九州農業試験場彙報, 1962(7): 419-495. HIDEO S. Study on lodging of rice[J]. Kyushu Agricultural Experiment Station Dictionary, 1962(7): 419-495. |
| [22] |
邱磊, 李艳欣, 张校晗, 魏海峰, 段佑强, 戴立国. 优质抗倒伏水稻新品种龙桦3特性及抗倒性状分析[J]. 种子科技, 2023, 41(16): 24-26. DOI:10.19904/j.cnki.cn14-1160/s.2023.16.007 QIU L, LI Y X, ZHANG X H, WEI H F, DUAN Y Q, DAI L G. Characteristics and lodging resistance analysis of high quality lodging resistant new rice varieties longhua 3[J]. Seed Science & Technology, 2023, 41(16): 24-26. DOI:10.19904/j.cnki.cn14-1160/s.2023.16.007 |
| [23] |
孙旭初. 水稻茎秆抗倒伏性的研究[J]. 中国农业科学, 1987(4): 32-37. SUN X C. Study on lodging resistance of rice stem[J]. Scientia Agricultura Sinica, 1987(4): 32-37. |
| [24] |
袁新捷, 刘潇, 陈国兴. 水稻核心种质资源茎秆抗倒伏性研究[J]. 华中农业大学学报, 2021, 40(1): 147-153. DOI:10.13300/j.cnki.hnlkxb.2021.01.018 YUAN X J, LIU X, CHEN G X. Stem lodging resistance of rice core germplasm[J]. Journal of Huazhong Agricultural University, 2021, 40(1): 147-153. DOI:10.13300/j.cnki.hnlkxb.2021.01.018 |
| [25] |
梁康迳, 杨仁崔, 杨蜀岚. 水稻茎秆性状的遗传效应分析[A]. 全国作物育种学术讨论会论文集[M]. 北京: 中国农业科学技术出版杜, 1998: 80-86. LIANG K J, YANG R C, YANG S L. Genetic effect analysis of stem traits in rice[A]. Proceedings of the national symposium on crop breeding[M]. Beijing: China Agriculture Science and Technology Press, 1998: 80-86. |
| [26] |
梁康迳, 王雪仁, 章清杞, 陈志雄. 基因型×环境互作效应对水稻茎秆抗倒性杂种优势的影响[J]. 福建农业大学学报, 2000(1): 12-17. DOI:10.13323/j.cnki.j.fafu(nat.sci.).2000.01.003 LIANG K J, WANG X R, ZHANG Q Q, CHEN Z X. Effect of genotype×environment interaction on heterosis for lodging resistance of the culm in cice[J]. Journal of Fujian Agricultural University, 2000(1): 12-17. DOI:10.13323/j.cnki.j.fafu(nat.sci.).2000.01.003 |
| [27] |
KEERTHIRAJ B, BIJU S. Genetic variability, heritability and genetic advance of yield and lodging-related traits in rice (Oryza sativa L.)[J]. Electronic Journal of Plant Breeding, 2020, 11(4): 1093-1098. DOI:10.37992/2020.1104.177 |
| [28] |
王晓飞, 陆展华, 刘维, 卢东柏, 王石光, 巫浩翔, 方志强, 何秀英. "绿色革命"以来水稻抗倒伏研究进展[J]. 广东农业科学, 2022, 49(3): 1-13. DOI:10.16768/j.issn.1004-874X.2022.03.001 WANG X F, LU Z H, LIU W, LU D B, WANG S G, WU H X, FANG Z Q, HE X Y. Advances in lodging resistance of rice since the "green revolution"[J]. Guangdong Agricultural Sciences, 2022, 49(3): 1-13. DOI:10.16768/j.issn.1004-874X.2022.03.001 |
| [29] |
LIU Q, YIN C S, LI X, HE C Q, DING Z, DU X. Lodging resistance of rice plants studied from the perspective of culm mechanical properties, carbon framework, free volume, and chemical composition[J]. Scientific Reports, 2022, 12(1): 20026. DOI:10.1038/s41598-022-24714-4 |
| [30] |
LUO X Y, WU Z F, FU L, DAN Z W, LONG W X, ZEHN Q Y, LIANG T, ZHU R S, HU Z L, WU X T. Responses of the lodging resistance of indica rice cultivars to temperature and solar radiation under field conditions[J]. Agronomy, 2022, 12(11): 2603. DOI:10.3390/agronomy12112603 |
| [31] |
何懿. 6个水稻品种的倒伏指数分析及抗倒伏性评价[J]. 农业科技通讯, 2023(6): 96-101, 106. HE Y. Lodging index analysis and lodging resistance evaluation of 6 rice varieties[J]. Bulletin of Agricultural Science and Technology, 2023(6): 96-101, 106. |
| [32] |
NOMURA T, ARAKAWA N, YAMAMOTO T, UEDA T, ADACHI S, YONEMARU J I, ABE A, TAKAGI H, YOKOYAMA T, OOKAWA T. Next generation gong-gulm rice with superior lodging resistance and high grain yield, monster rice 1[J]. PLoS One, 2019, 14(8): e0221424. DOI:10.1371/journal.pone.0221424 |
| [33] |
WANG X Y, XU L, LI X X, YANG G D, WANG F, PENG S B. Grain yield and lodging-related traits of ultrashort-duration varieties for direct-seeded and double-season rice in central china[J]. Journal of Integrative Agriculture, 2022, 21(10): 2888-2899. DOI:10.1016/j.jia.2022.07.035 |
| [34] |
TSUGAWA S, SHIMA H, ISHIMOTO Y, ISHIKAWA K. Thickness-stiffness trade-off improves lodging resistance in rice[J]. Scientific Reports, 2023, 13(1): 10828. DOI:10.1038/s41598-023-37992-3 |
| [35] |
FAN C F, LI Y, HU Z, HU H Z, WANG G Y, LI A, WANG Y M, TU Y Y, XIA T, PENG L C, FENG S Q. Ectopic expression of a novel osextensin-like gene consistently enhances plant lodging resistance by regulating cell elongation and cell wall thickening in rice[J]. Plant Biotechnology Journal, 2018, 16(1): 254-263. DOI:10.1111/pbi.12766 |
| [36] |
OLAGUNJU S O, ATAYESE M O, SAKARIYAWO O S, DARE E O, NASSIR A L. Culm morphological traits contributing to lodging resistance in first generation nerica cultivars under foliar application of orthosilicic aid fertilizer[J]. Silicon, 2021, 13: 3059-3073. DOI:10.1007/s12633-020-00652-6 |
| [37] |
WU D, WU D, FENG H, NIU Z Y, XIONG L Z, YANG W N. A deep learning-integrated micro-ct image analysis pipeline for quantifying rice lodging resistance-related traits[J]. Plant Communications, 2021, 2(2): 1-12. DOI:10.1016/j.xplc.2021.100165 |
| [38] |
MIAO W, LI F C, LU J C, WANG D L, CHEN M K, TANG L, XU Z J, CHEN W F. Biochar application enhanced rice biomass production and lodging resistance via promoting co-deposition of silica with hemicellulose and lignin[J]. Science of the Total Environment, 2023, 855: 158818. DOI:10.1016/j.scitotenv.2022.158818 |
| [39] |
LI Q, FU C F, LIANG C L, NI X J, ZHAO X Z, CHEN M, QU L J. Crop lodging and the roles of lignin, cellulose, and hemicellulose in lodging resistance[J]. Agronomy, 2022, 12(8): 1795. DOI:10.3390/agronomy12081795 |
(责任编辑 张辉玲)