 2023, Vol. 50
2023, Vol. 50文章信息
引用本文 |

基金项目
- 清远市科技计划项目(DZXQY017);科技创新战略专项(高水平农科院建设)(202110TD,R2020PY-JC001);广东省农业科学院“十四五”学科团队项目(202122TD)
作者简介
- 连聪(1998—),女,在读硕士生,研究方向为动物疫病诊断与防控,E-mail:3326906588@qq.com.
通讯作者
- 罗胜军(1974—),男,硕士,副研究员,研究方向为动物疫病诊断,E-mail:422368881@qq.com.
文章历史
- 收稿日期:2022-07-06




新城疫(Newcastle Disease,ND)是一种由新城疫病毒(Newcastle Disease Virus,NDV)引起的禽类急性、接触性传染病[1],是严重损害养禽业发展的疾病之一,世界动物卫生组织将其划为A类传染病,我国将其列为一类传染病。自然条件下,NDV经受伤皮肤、消化道、呼吸道黏膜等部位侵入机体,也可通过垂直传播导致幼禽感染。新城疫可引起鸡、鸭、鹅、鸽子、鹌鹑等禽类感染,尤其鸡受感染后通常会导致死亡,其他动物的症状一般比鸡轻[2]。ND一年四季均可发生,春秋季多发,临床上可根据发病速度、症状及死亡率将其分为最急性、急性、慢性3种类型,其中急性ND普遍流行,死亡率高(可达90% 以上)。NDV存在于病禽的所有组织器官、体液及分泌物中,脑、脾、肺含毒量最高,故病禽会出现转脖、站立不稳等神经症状,脾脏充血、出血,亦可见明显的呼吸困难、咳嗽和气喘等呼吸道症状[3]。
ND自1926年在印度尼西亚被首次发现,至今已引发4次全球性大规模流行[4],从病毒流行史上看,ND一直是家禽的毁灭性疾病,目前该疾病在许多国家仍然是影响家禽养殖业发展的主要问题之一。即使在ND已经得到控制的国家,依然有较重的经济负担,主要用于疫苗接种和维持严格的生物安全措施[5]。目前市场上存在多种活疫苗和灭活疫苗,包括易于保存、运输的耐热NDV疫苗[6],国内普遍使用灭活疫苗防控ND。ND经常被误诊为沙门氏菌病、螺旋体病、喉气管炎和某些出血性疾病,故对其诊断需要密切的监测,以迅速、特异性地识别病原。血清学检测和分子生物学检测是NDV检测的标准方法,血清学检测可短时间内检测到NDV的存在,从而可迅速采取行动控制疫情传播;分子生物学检测则确定其基因分型,为后续疫苗研发提供方向。血清学检测主要采用血凝抑制试验与ELISA检测。血凝抑制试验主要测量NDV特异性抗体抑制NDV颗粒对红细胞凝集的能力,因其成本较低及操作简便而成为目前主要的检测方法。ELISA检测高度敏感,产生的结果与血凝抑制试验结果基本一致,也是常用的检测方法。近年来,分子生物学检测方法迅速发展,各新型技术的研发取得了重大成果,其中微阵列杂交技术、生物传感器和下一代测序最受关注,但因成本高、技术复杂等原因尚未被广泛使用,而普遍使用的技术是逆转录聚合酶链式反应(RT-PCR)和荧光定量聚合酶链式反应(qPCR)。qPCR技术使用最广,相较于传统检测技术,该技术更便捷且具有更高的病毒检测灵敏度。分子生物学技术的进步对分子流行病学的研究提供了有利条件,本文就NDV流行病学及其基因型的遗传进化等方面研究进展进行综述,以期为NDV的防控和疫苗研发提供参考。
1 NDV病原学NDV对外界抵抗力较强,在自然环境中可生存较长时间;对低温抵抗力强,低温下可存活数月;60 ℃环境下经45 min可灭活,紫外线照射30 min被灭活[7];对乙醚等有机溶剂敏感。NDV是单股负链RNA病毒,为副黏病毒科新城疫样病毒属的禽副黏病毒Ⅰ型[8]。NDV多为圆形,直径100~250 nm,有囊膜,单链不分节。NDV基因组有3种长度,分别为15 186、15 192、15 198 nt[9],结构为3'-NP-P-M-F-HN-L-5'(图 1),编码6种特异性结构蛋白[10],分别为核衣壳蛋白(NP)、磷蛋白(P)、基质蛋白(M)、融合蛋白(F)、血凝素-神经氨酸酶蛋白(HN)和大蛋白(L)。每个特异性结构蛋白的基因都存在起始因子和终止因子序列,各蛋白基因之间通过基因间序列将彼此分隔开,保证其表达的准确性。

1.1 血凝素-神经氨酸酶蛋白(HN)
HN蛋白和F蛋白为刺突糖蛋白,刺突长度约为8 nm,分别以三聚体和四聚体的形式存在,位于囊膜表面,在病毒感染过程中参与病毒的入侵,可诱导保护性免疫[11],它们均为病毒热稳定性的决定因素[12]。HN是一种Ⅱ型整合膜蛋白,蛋白分子量为74 kD,可使病毒吸附在细胞表面的唾液酸受体,并通过血凝素、神经氨酸酶的生物学活性破坏受体功能,参与病毒入侵。HN的受体识别位点和神经氨酸酶活性位点均位于球状头部,并且高度保守,球状头部区域也被认为是抗体结合位点[13]。由于终止密码子位置不同,自然界中存在不同长度的HN蛋白,最短的HN蛋白含571个氨基酸,存在于速发菌株中;最长的HN蛋白含616个氨基酸,存在于缓发菌株中。HN蛋白中扩展羧基末端的长度和序列可影响蛋白功能,但尚未明确其对NDV毒力的作用[5]。
1.2 融合蛋白(F)F蛋白是存在于NDV包膜上的表面糖蛋白,参与病毒穿入、细胞融合等过程。其基因组序列用于对NDV毒株基因型进行分类,切割位点处序列是世界动物卫生组织(OIE)公认的毒力指标。NDV系统性传播及其毒力大小取决于F蛋白在组织中的裂解。根据OIE相关标准,位于112~116位氨基酸之间的F蛋白切割位点具有多个碱性氨基酸残基,并在117位具有苯丙氨酸残基的毒株被认为是强毒株;在117位具有一元F切割位点和亮氨酸残基的毒株被认为是弱毒株[2]。
1.3 基质蛋白(M)M蛋白本质为疏水蛋白,位于核衣壳和脂质膜之间,分子量约为40 kD,由364个氨基酸组成。M蛋白是一种碱性蛋白质,具有与病毒核酸相互作用的区域(17 bp),包括9个碱性氨基酸[14],其在RNA合成及病毒自身装配中起关键作用[15]。M蛋白在副粘病毒中高度保守,群体发生突变后其存在极少的非同义碱基替代,这可作为对不同地域NDV分离株进行分类的依据[16]。
1.4 大蛋白(L)L蛋白是NDV基因组中最大的蛋白质,由2 204个氨基酸组成,分子量为250 kD[9]。L蛋白属于RNA依赖性RNA聚合酶,与病毒组装、合成相关,在病毒感染过程中充当病毒复制酶和转录酶,其合成病毒mRNA并协助基因组RNA复制,还对新形成的mRNA进行5' 端加帽、甲基化及激活多聚A聚合酶活性[17]。L蛋白可调节NDV的毒力,可能通过增加复制期间病毒RNA的合成速率而发挥有效作用[18]。
1.5 磷蛋白(P)P蛋白连接L蛋白与NP蛋白,是聚合酶的辅助因子。P蛋白由395个氨基酸组成,在特定的丝氨酸和苏氨酸残基处被磷酸化,并作为同源寡聚体发挥作用,其在病毒复制和转录中也具有至关重要的作用[5]。P蛋白的四聚体介导L蛋白和N-RNA模板之间的相互作用,以防止NP蛋白随机包裹非病毒RNA。此外,P蛋白与未组装的NP蛋白形成复合物,调节逆转录过程。在病毒复制过程中,P蛋白的不同结构域与NP蛋白相互作用时发挥不同功能。NDV的6个结构蛋白基因中有5个蛋白基因编码单一的蛋白质,只有P蛋白基因通过RNA编辑编码3个蛋白质,即P蛋白、V蛋白和W蛋白[9]。V蛋白和W蛋白是辅助蛋白,仅存在于病毒感染的细胞中,其中V蛋白属于干扰素(IFN)拮抗剂,对NDV毒力大小起重要作用[19]。
1.6 核衣壳蛋白(NP)NP蛋白含489个氨基酸,分子量为55 kD,覆盖整个病毒核酸形成核糖核蛋白(RNP),以保护RNA免受核酸酶的侵害[16]。NP蛋白是病毒颗粒中最丰富的蛋白质,电子显微镜下可见“人”字形结构,是病毒复制和mRNA生物合成所需的最小模板,与N蛋白、P蛋白、L蛋白及基因组RNA结合形成RNP。
2 NDV分子流行病学进展 2.1 NDV基因型F蛋白是主要的特异性结构蛋白,根据其核苷酸序列划分NDV的基因型,其基因全长1 792 nt,ORF长1 662 nt,编码553个氨基酸,分子量约55 kD[20-21]。根据其基因编码可将NDV分为I和Ⅱ两类。若F蛋白裂解位点的氨基酸组成为112R/K-R-Q-K/R-R-F117,则其为I类毒株;若裂解位点的氨基酸组成为112G/EK/R-Q-G/ E-R-L117,则其为Ⅱ类毒株[22]。I类毒株主要来源于野生鸟类,大多毒株为低毒力[23],家禽中主要感染鸭、鹅等水禽,鸡群中偶尔出现,但概率较小。Ⅱ类毒株是可引起禽类明显症状的强毒株,具有多种基因型,分为I型、Ⅱ型、Ⅲ型等,遗传进化距离大于0.1就被判定为不同基因型[3]。巴基斯坦和孟加拉国曾从鸽子中分离出基因型XXI.1.2毒株[24-25]。因此,Ⅱ类毒株至少分为Ⅰ~XXI达20多种。Ⅱ类毒株在野生鸟类中具有某些特定的基因型,例如基因型Ⅵ型毒株为鸽子、鸬鹚等野生鸟类的特有毒株[26],而至今并无证据表明该型毒株无感染家禽的风险。
我国曾分离出多种基因型NDV毒株。1946年,我国分离到特有的Ⅱ类Ⅸ基因型F48E9毒株[27],为嗜神经型强毒株,家禽表现出以神经症状为主导的临床特征[28],即出现翅麻痹、跛行、站立不稳、头颈向后侧扭转、伏地旋转等症状;在新城疫第3次大流行时,主要流行株为Ⅱ类Ⅵ型毒株;20世纪90年代NDV的Ⅱ类Ⅶ d型成为我国主要优势基因型[29];2010年,广东首次监测到Ⅱ类XII型,并证实为强毒株[30]。这些毒株的成功分离为新城疫的诊断与防控提供了巨大帮助。进入21世纪以来,NDV Ⅰ类毒株的致病性有增强趋势,从而引起人们的关注,2008年首次分离出Ⅰ类病毒Duck China/08-004/2008[31],自此逐渐加强对NDV Ⅰ类病毒的研究,而高致病性的Ⅱ类毒株始终是人们研究新城疫疾病的重点。到目前为止,NDV的基因型仍在不断改变,掌握其所有基因型并研究有针对性的防治方法才有可能净化NDV。
2.2 分子流行病学2.2.1 我国NDV不同基因型流行情况 为了解我国部分地区NDV流行毒株的基因型情况,本文统计整理了中国知网和PubMed报道的NDV分子流行病学数据,结果(表 1)发现,2008—2020年间在各地区收集的14 062份样本中共分离到234株毒株,其中166株鉴定出具体基因型,涉及基因型Ⅰ类1型(4/166,2.41%)、2型(3/166,1.81%)、3型(60/166,36.14%),基因型Ⅱ类Ⅰ型(26/166,15.66%)、Ⅱ型(23/166,13.86%)、Ⅵ型(2/166,1.2%)、Ⅶ型(37/166,22.29%)、Ⅷ型(3/166,1.81%)、Ⅸ(7/166,4.22%)、Ⅻ(1/166,0.6%)。
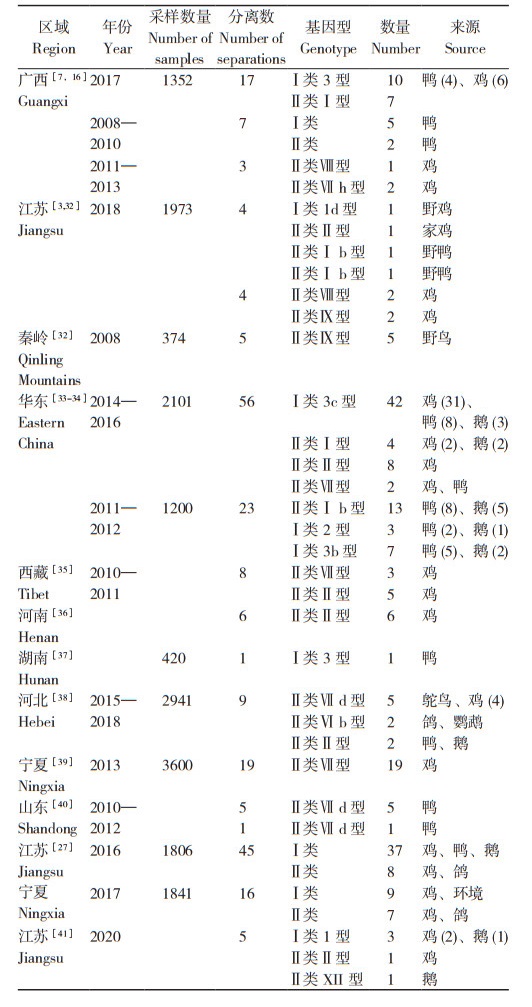
由图 2可知,NDV在鸡流行毒株中,Ⅰ类3型(37/93,39.78%)占比最高,其次为Ⅱ类Ⅶ型(27/93,29.03%),Ⅱ类Ⅱ型(21/93,22.58%)则占比较少;在鸭流行毒株中,Ⅰ类3型(18/43,41.86%)占比最高,其次为Ⅱ类Ⅰ型(15/43,34.88%),Ⅱ类Ⅶ(7/43,16.28%)占比较低;在鹅流行毒株中,Ⅱ类Ⅰ型(7/16,43.75%)占比最高,其次为Ⅰ类3型(5/16,31.25%)、Ⅰ类1型(1/16,6.25%),此外还发现了1株Ⅱ类Ⅻ型毒株,未发现Ⅱ类Ⅶ型病毒,推测其可能在鹅中感染率不高。Ⅰ类3型毒株在鸡、鸭、鹅中均有较强感染力,为保证家禽健康,加强对该型毒株研究具有重大意义。

|
| 图 2 不同家禽的NDV毒株基因型统计 Fig. 2 Genotype statistics of NDV strains in different poultry species |
2.2.2 遗传进化分析 在我国分离出的NDV多种基因型毒株中,Ⅱ类Ⅸ型是我国特有的毒株,而Ⅱ类Ⅶ型病毒现已成为我国NDV流行的主要优势基因型[24]。我国西藏、广西、河北、宁夏及山东、江苏等华东地区都曾分离出Ⅱ类Ⅶ型毒株,其中山东、江苏、河北分离到Ⅶ d亚型。由此表明,在基因Ⅶ型毒株中,Ⅶ d亚型更为流行。
ND感染病例中,鸡感染最多,以雏鸡尤甚。其影响鸡的消化系统、呼吸系统、神经系统,有时亦影响生殖系统。疾病表现受个体的易感性、病毒依赖性及其他压力因素的影响。感染后所呈现的症状因基因型不同而异。感染Ⅰ类中的弱毒株,多表现为轻型症状,甚至不引起任何临床症状。若感染Ⅱ类中的强毒株,则会引起机体明显的临床症状,甚至导致大批量死亡,故Ⅱ类毒株是研究和预防的主要方向。F蛋白切割位点是病毒毒力的决定因素。本文选取了NCBI中15株不同基因型NDV的F基因序列,用MEGAX软件的最大似然法对这些毒株的F基因序列进行分析,构建了基于F基因序列的系统发育树(图 3)。当前,基因Ⅱ型与基因Ⅶ型NDV在鸡群中较为流行,根据系统发育树可知,基因Ⅰ型、Ⅱ型、Ⅸ型与基因Ⅶ型、Ⅷ型相距甚远,针对基因Ⅱ型毒株的疫苗对基因Ⅶ型毒株的预防效果不佳,因此,在疫苗接种时,结合当地及养殖场内实际情况,应选择最完善的接种方案。

|
| 图 3 NDV基于F基因的系统遗传进化树 Fig. 3 Phylogenetic evolutionary tree of NDV based on F gene |
将Ⅱ类Ⅰ型、Ⅱ型、Ⅶ型、Ⅷ型和Ⅸ型毒株的核苷酸序列进行比对(图 4),Ⅰ型病毒之间同源性为90.7%~93.8%,与其他基因型病毒的同源性为85.6%~99.8%;Ⅱ型病毒之间同源性为89.5%~99.6%,与其他基因型病毒的同源性为85.1%~99.8%;Ⅶ型病毒之间同源性为85.2%~99.1%,与其他病毒的同源性为84.5%~95.6%;Ⅷ型病毒之间同源性为95.3%,与其他病毒的同源性为84.0%~94.4%;Ⅸ型病毒之间同源性为84.8%~95.6%,与其他病毒之间的同源性为84.0%~91.3%。各基因型病毒之间同源性相差不大,相比之下,Ⅶ型与Ⅷ型病毒与其他病毒之间同源性较小,Ⅸ型病毒与其他毒株同源性更小。

|
| 1: Ⅰ-M24693; 2: Ⅰ-KX352834; 3: Ⅰ-HG326605; 4: Ⅱ -FJ705468; 5: Ⅱ - FJ705466; 6: Ⅱ - KX857721; 7: Ⅶ - JN986837;8: Ⅶ - KX268351; 9: Ⅶ - HQ697254; 10: Ⅶ - AB853927; 11: Ⅷ - JX012096; 12: Ⅷ - FJ751918; 13: Ⅸ - FJ436302; 14: Ⅸ - FJ436303; 15: Ⅸ - AF458009 图 4 NDV中F基因核苷酸序列同源性分析结果 Fig. 4 Results of nucleotide sequence homology analysis of F gene in NDV |
3 我国新城疫流行情况与免疫防控
为了解我国部分地区新城疫流行情况,本文整理2010—2020年间的相关文献,得到不同省区的NDV阳性情况。由表 2可知,抗体检测共45534份,阳性样本41698份,阳性率为91.58%;抗原检测共37 568份,阳性样本789份,阳性率为2.10%。抗体检测显示疫苗接种率良好,各地区间抗原阳性率在11.00% 以下,具有较大差异,在具有NDV疫情的地区需结合该地区的流行毒株来筛选疫苗,从而更科学、精准地防控NDV。

我国新城疫目前多为地方性散发流行,NDV中的外源基因具有稳定性[51],故疫苗的广泛使用可有效减少NDV的传播,使疾病发生率显著下降。目前,常见新城疫疫苗主要为B1、LaSota和V4等毒株的减毒活疫苗[52],以及LaSota和VH等毒株的灭活疫苗[53]。疫苗是预防NDV发生、传播的有效手段,但由于NDV的F基因具有较高变异率,基因型种类也不断增多,疫苗免疫后并不能完全阻止流行毒株的侵袭[54]。因此,临床上NDV防控中疫苗(种类和基因型)的选择至关重要,免疫方案的制定需要结合本地NDV的实际流行情况,灭活疫苗和减毒活疫苗联合使用有助于NDV阳性场的控制,而选择与NDV流行毒株同源性较高的疫苗株进行免疫,对本场NDV的净化具有重要意义。此外,我们依然要做好传染源的防控,定期对场内环境进行检疫,提高人员防疫意识,从根源上预防疾病暴发与传播[55]。
Ⅰ类NDV主要分离自野生鸟类,且大部分是低毒力甚至无毒力的毒株[56],因此,我国NDV的防控措施主要针对Ⅱ类毒株。若能加强对Ⅱ类Ⅱ型和Ⅶ型这两种主要流行毒株的研究,则能使防控工作取得更好成效。掌握主要感染的NDV基因型有助于了解疾病的流行特征,这为我国NDV防控和新型疫苗研发提供依据。
4 结语本文通过文献对我国不同地区NDV流行情况进行统计,发现NDV主要分布于华南地区、华东地区及部分西北地区,各地区暴发程度具有较大差异,候鸟迁徙区域感染更为明显。2010—2020年间,感染较严重地区的阳性率在11.0% 左右,有些地区无阳性记录,针对NDV防控需结合当地实际情况制定策略。对流行地区的NDV基因型进行整理,发现Ⅰ类3型、Ⅱ类Ⅱ型和Ⅱ类Ⅶ型是我国鸡群中NDV的主要流行基因型,Ⅰ类3型和Ⅱ类Ⅰ型基因型是水禽(如鸭、鹅)的主要流行株。Ⅰ类毒株为弱毒株,临床表现症状较轻,近年来其症状有加重趋势,而目前对NDV的研究依然以Ⅱ类毒株为主。Ⅰ类3型毒株对鸡、鸭、鹅均有较强感染性,且其更易通过野生鸟类进行传播,故加强对Ⅰ类毒株的研究很有必要。通过对不同Ⅱ类毒株的F基因序列进行比对,发现Ⅱ类Ⅰ型毒株与Ⅱ型毒株间同源性为89.7%~99.8%,Ⅰ型毒株与Ⅶ型毒株间同源性为86.6%~90.9%,Ⅱ型毒株与Ⅶ型毒株间同源性为85.4%~95.6%。
ND是全球家禽业的主要威胁因素,不同程度毒力的NDV毒株在禽类中广泛传播。ND与其他疾病相比传播率更高,传播范围更广。卫生条件差、营养缺乏、疫苗接种不完善、与其他禽类接触等均能促进ND的传播。在疾病暴发前切断可能感染的途径,进行免疫保护是预防ND的有效措施[57]。加强对病毒的监测,即使对Ⅰ类中的弱毒株也不能掉以轻心,病毒入侵可使机体防御力显著降低,极可能发生继发性感染。继发性细菌感染是病毒感染的常见后遗症,相比轻型NDV更具有破坏性。不断完善防御策略是控制NDV传播的必要手段,由于NDV传播途径多样,野生鸟类、商业鸟类、宠物鸟类均有病毒传播风险,人们以往对其并无过多关注,这是ND预防策略中较为薄弱的点,加强对这些鸟类的控制,严格依照生物安全标准进行饲养、检疫,加固对NDV的防御墙。一种优秀的NDV疫苗可以有效预防临床疾病,目前可用的灭活和减毒活疫苗只能预防相应的病毒感染,不能避免异源病毒入侵。研制新型疫苗及优化防治策略的工作仍须继续,以期在不久的将来找到有针对性、更简捷精确控制NDV的方法。
| [1] |
李晓, 杨兴艳. NDV疫苗核心专利技术分析[J]. 中国科技信息, 2022(7): 23-25. LI X, YANG X Y. Analysis of the core patent technology of NDV vaccine[J]. China Science and Technology Information, 2022(7): 23-25. |
| [2] |
GAO P, CHEN L, FAN L, REN J, DU H, SUN M, LI Y, XIE P, LIN Q, LIAO M, XU C, NING Z, DING C, XIANG B, REN T. Newcastle disease virus RNA-induced IL-1β expression via the NLRP3/caspase-1 inflammasome[J]. Veterinary Research, 2020, 51(1): 53. DOI:10.1186/s13567-020-00774-0 |
| [3] |
BELLO M B, YUSOFF K, IDERIS A, HAIR-BEJO M, PEETERS B P H, OMAR A R. Diagnostic and vaccination approaches for Newcastle Disease Virus in poultry: The current and emerging perspectives[J]. Biomed Research International, 2018, 2018: 1-18. DOI:10.1155/2018/7278459 |
| [4] |
陈林娜, 孙军峰, 李乐, 师乾凯, 邸涛, 刘添仪, 赵冉, 张春玮, 韩宗玺, 刘胜旺. 基因Ⅶ型新城疫病毒融合前构象F蛋白的表达及结构分析[J]. 中国兽医科学, 2021, 51(4): 475-482. DOI:10.16656/j.issn.1673-4696.2021.0065 CHEN L N, SUN J F, LI L, SHI Q K, DI T, LIU T Y, ZHAO R, ZHANG C W, HAN Z X, LIU S W. Expression and structural analysis of the pre-fusion conformational F protein of genotype Ⅶ Newcastle disease virus[J]. Chinese Veterinary Science, 2021, 51(4): 475-482. DOI:10.16656/j.issn.1673-4696.2021.0065 |
| [5] |
ALEXANDER D J. Newcastle disease and other avian paramyxoviruses[J]. Revue Scientifique et Technique (International Office of Epizootics), 2000, 19(2): 443-462. DOI:10.20506/rst.19.2.1231 |
| [6] |
GANAR K, DAS M, SINHA S, KUMAR S. Newcastle disease virus: current status and our understanding[J]. Virus Research, 2014, 184: 71-81. DOI:10.1016/j.virusres.2014.02.016 |
| [7] |
翟国胜. NDV广西分离株分离鉴定、基因序列分析及qRT-PCR检测方法的建立[D]. 南宁: 广西大学, 2015. DOI: 10.27034/d.cnki.ggxiu.2015.000039. ZHAI G S. Isolation and identification of NDV Guangxi isolates, gene sequence analysis and establishment of qRT-PCR detection method[D]. Nanning: Guangxi University, 2015. DOI: 10.27034/d.cnki.ggxiu.2015.000039. |
| [8] |
张振兴, 郭亮, 宋建领. 鸡新城疫流行原因分析及防控[J]. 畜牧兽医科技信息, 2022(2): 156-158. DOI:10.3969/J.ISSN.1671-6027.2022.02.067 ZHANG Z X, GUO L, SONG J L. Analysis of epidemic causes and prevention and control of Newcastle disease in chickens[J]. Animal Husbandry and Veterinary Science and Technology Information, 2022(2): 156-158. DOI:10.3969/J.ISSN.1671-6027.2022.02.067 |
| [9] |
CZEGLÉDI A, UJVÁRI D, SOMOGYI E, WEHMANN E, WERNER O, LOMNICZI B. Third genome size category of avian paramyxovirus serotype 1 (Newcastle disease virus) and evolutionary implications[J]. Virus Research, 2006, 120(1/2): 36-48. DOI:10.1016/j.virusres.2005.11.009 |
| [10] |
邓珊珊. 鸡基质蛋白3在新城疫病毒复制中的作用研究[D]. 贵阳: 贵州大学, 2019. DENG S S. Study on the role of chicken matrix protein 3 in Newcastle disease virus replication[D]. Guiyang: Guizhou University, 2019. |
| [11] |
YUSOFF K, TAN W S. Newcastle disease virus: macromolecules and opportunities[J]. Avian Pathology, 2001, 30(5): 439-455. DOI:10.1080/03079450120078626 |
| [12] |
LIU T, SONG Y, YANG Y, BU Y, CHENG J, ZHANG G, XUE J. Hemagglutinin-Neuraminidase and fusion genes are determinants of NDV thermostability[J]. Veterinary Microbiology, 2019, 228: 53-60. DOI:10.1016/j.vetmic.2018.11.013 |
| [13] |
IORIO R M, MAHON P J. Paramyxoviruses: different receptors-different mechanisms of fusion[J]. Trends in Microbiology, 2008, 16(4): 135-137. DOI:10.1016/j.tim.2008.01.006 |
| [14] |
CHAMBERS P, MILLAR N S, PLATT S G, EMMERSON P T. Nucleotide sequence of the gene encoding the matrix protein of Newcast le disease vir us[J]. Nucleic Acids Research, 1986, 14(22): 9051-9061. DOI:10.1093/nar/14.22.9051 |
| [15] |
BATTISTI A J, MENG G, WINKLER DC, MCGINNES L W, PLEVKA P, STEVEN A C, MORRISON T G, ROSSMANN M G. Structure and assembly of a paramyxovirus matrix protein[J]. Proceedings of the National Acadamy of Sciences of the United States of America, 2012, 109(35): 13996-14000. DOI:10.1073/pnas.1210275109 |
| [16] |
SEAL B S, KING D J, MEINERSMANN R J. Molecular evolution of the Newcastle disease virus matrix protein gene and phylogenetic relationships among the paramyxoviridae[J]. Virus Research, 2000, 66(1): 1-11. DOI:10.1016/s0168-1702(99)00119-7 |
| [17] |
DORTMANS J C, ROTTIER P J, KOCH G, PEETERS B P. The viral replication complex is associated with the virulence of Newcastle disease virus[J]. Journal of Virology, 2010, 84(19): 10113-10120. DOI:10.1128/JVI.00097-10 |
| [18] |
ROUT S N, SAMAL S K. The large polymerase protein is associated with the virulence of Newcastle disease virus[J]. Journal of Virology, 2008, 82(16): 7828-36. DOI:10.1128/JVI.00578-08 |
| [19] |
MENG Q, HE J, ZHONG L, ZHAO Y. Advances in the study of antitumour immunotherapy for Newcastle disease virus[J]. International Journal of Medical Sciences, 2021, 18(11): 2294-2302. DOI:10.7150/ijms.59185 |
| [20] |
EBRAHIMI M M, SHAHSAVANDI S, MOAZENIJULA G, SHAMSARA M. Phylogeny and evolution of Newcastle disease virus genotypes isolated in Asia during 2008-2011[J]. Virus Genes, 2012, 45(1): 63-68. DOI:10.1007/s11262-012-0738-5 |
| [21] |
陈礼朋. 鸡新城疫病毒HNO9-69株主要基因的克隆、原核表达及鉴定[D]. 郑州: 河南农业大学, 2012. CHEN L P. Cloning, prokaryotic expression and identification of major genes of chicken Newcastle disease virus strain HNO9-69[D]. Zhengzhou: Henan Agricultural University, 2012. |
| [22] |
魏潇. 表达新城疫病毒F蛋白传染性喉气管炎病毒gC缺失株的构建及评价[D]. 北京: 中国农业科学院, 2020. DOI: 10.27630/d.cnki.gznky.2020.000288. WEI X. Construction and evaluation of an infectious laryngotracheitis virus gC-deficient strain expressing Newcastle disease virus F protein[D]. Beijing: Chinese Academy of Agricultural Sciences, 2020. DOI: 10.27630/d.cnki.gznky.2020.000288. |
| [23] |
ABSALON A E, CORTÉS-ESPINOSA D V, LUCIO E, MILLER P J, AFONSO C L. Epidemiology, control, and prevention of Newcastle disease in endemic regions: Latin America[J]. Tropical Animal Health and Production, 2019, 51(5): 1033-1048. DOI:10.1007/s11250-019-01843-z |
| [24] |
NOORUZZAMAN M, BARMAN L R, MUMU T T, CHOWDHURY E H, DIMITROV K M, ISLAM M R. A Pigeon-Derived Sub-Genotype XXI.1.2 Newcastle Disease Virus from Bangladesh Induces High Mortality in Chickens[J]. Viruses, 2021, 13(8): 1520. DOI:10.3390/v13081520 |
| [25] |
NASIR S, WAJID A, NAUREEN A, MUSTAFA A, AYUB G, AIN Q, DIN A M, BATOOL A, HUSSAIN T. Isolation and phylogenetic analysis of Avian orthoavulavirus 1 sub-genotypes VⅡ.2 and XXI.1.2 from caged birds in the Lahore district, Pakistan-Short communication[J]. Acta Veterinaria Hungarica, 2021, 70(1): 73-76. DOI:10.1556/004.2021.00053 |
| [26] |
DIEL D G, MILLER P J, WOLF P C, MICKLEY R M, MUSANTE A R, EMANUELI D C, SHIVELY K J, PEDERSEN K, AFONSO C L. Characterization of Newcastle disease viruses isolated from cormorant and gull species in the United States in 2010[J]. Avian Diseases, 2012, 56: 128-133. DOI:10.1637/9886-081111-Reg.1 |
| [27] |
高晓敏. 2018-2020年我国新城疫病毒分子流行病学及针对流行毒株的新疫苗研制[D]. 扬州: 扬州大学, 2021. DOI: 10.27441/d.cnki.gyzdu.2021.002258. GAO X M. Molecular epidemiology of Newcastle disease virus and development of new vaccines against prevalent strains in China in 2018-2020[D]. Yangzhou: Yangzhou University, 2021. DOI: 10.27441/d.cnki.gyzdu.2021.002258. |
| [28] |
毕友坤, 屈阳, 卫巧林, 王文彬, 赵杰, 金忠元, 张淑霞, 王兴龙, 杨增岐, 萧飒. 新城疫病毒强毒株F48E9致病性细化分型的鉴定[J]. 畜牧与兽医, 2018, 50(2): 91-96. BI Y K, QU Y, WEI Q L, WANG W B, ZHAO J, JIN Z Y, ZHANG S X, WANG X L, YANG Z Q, XIAO S. Identification of a strong virulent strain of Newcastle disease virus F48E9 with refined pathogenicity typing[J]. Livestock and Veterinary Medicine, 2018, 50(2): 91-96. |
| [29] |
刘秀梵, 王志亮. 新城疫[M]. 北京: 中国农业出版社, 2015. LIU X F, WANG Z L. Newcastle disease[M]. Beijing: China Agricultural Press, 2015. |
| [30] |
谢丽基, 谢芝勋, 罗思思, 邓显文, 谢志勤, 王盛, 黄娇玲, 曾婷婷, 张艳芳, 黄莉, 范晴, 张民秀. 2017年广西地区活禽市场新城疫病原学监测与遗传进化分析[J]. 中国预防兽医学报, 2018, 40(8): 747-750. XIE L J, XIE Z X, LUO S S, DENG X W, XIE Z Q, WANG S, HUANG J L, ZENG T T, ZHANG Y F, HUANG L, FAN Q, ZHANG M X. Pathogenetic surveillance and genetic evolution analysis of Newcastle disease in live poultry markets in Guangxi region in 2017[J]. Chinese Journal of Preventive Veterinary Medicine, 2018, 40(8): 747-750. |
| [31] |
刘华雷, 蒋小刚, 张维, 赵云玲, 郑东霞, 孙承英, 陈飞, 王志亮. 一株Class I新城疫病毒中国分离株分子特性的研究[J]. 中国动物检疫, 2008, 25(8): 30-33. LIU H L, JIANG X G, ZHANG W, ZHAO Y L, ZHENG D X, SUN C Y, CHEN F, WANG Z L. Molecular characterization of a Chinese isolate of Class I Newcastle disease virus[J]. China Animal Quarantine, 2008, 25(8): 30-33. |
| [32] |
段旭基, 张鹏, 吴朋朋, 马静, 陈胜利, 郝华芳, 张定全, 韩青松, 付向晶, 张渭东, 杜恩岐, 杨增岐. 秦岭北麓关中地区野鸟源基因Ⅸ型新城疫病毒病原学监测及分离株毒力测定[J]. 中国兽医学报, 2013, 33(8): 1163-1169. DOI:10.16303/j.cnki.1005-4545.2013.08.003 DUAN X J, ZHANG P, WU P P, MA J, CHEN S L, HAO H F, ZHANG D Q, HAN Q S, FU X J, ZHANG W D, DU E Q, YANG Z Q. Pathogenetic surveillance and virulence determination of wild bird-derived genotype Ⅸ Newcastle disease virus isolates in Guanzhong area at the northern foot of Qinling Mountains[J]. Chinese Journal of Veterinary Medicine, 2013, 33(8): 1163-1169. DOI:10.16303/j.cnki.1005-4545.2013.08.003 |
| [33] |
刘靖. 2014-2016年华东地区新城疫病毒的分离鉴定及遗传进化分析[D]. 扬州: 扬州大学, 2017. LIU J. Isolation and identification of Newcastle disease virus and genetic evolutionary analysis in East China from 2014-2016[D]. Yangzhou: Yangzhou University, 2017. |
| [34] |
王珏, 仇旭升, 戴亚斌, 于圣青, 丁铲, 秦爱建. 华东地区健康水禽携带新城疫病毒情况调查[J]. 中国动物传染病学报, 2013, 21(6): 19-25. WANG J, QIU X S, DAI Y B, YU S Q, DING C, QIN A J. Survey of Newcastle disease virus in healthy waterfowl in East China[J]. Chinese Journal of Animal Infectious Diseases, 2013, 21(6): 19-25. |
| [35] |
朱洪云, 旦巴次仁. 藏鸡NDV的分离鉴定及F基因的遗传进化分析[J]. 西北农林科技大学学报(自然科学版), 2012, 40(10): 31-37. DOI:10.13207/j.cnki.jnwafu.2012.10.005 ZHU H Y, DANBA C R. Isolation and identification of NDV in Tibetan chickens and genetic evolutionary analysis of F gene[J]. Journal of Northwest Agriculture and Forestry University of Science and Technology (Natural Science Edition, 2012, 40(10): 31-37. DOI:10.13207/j.cnki.jnwafu.2012.10.005 |
| [36] |
阮涛. 河南部分地区NDV分子流行病学研究及NDV HN主要抗原区蛋白在抗体检测中的应用[D]. 新乡: 河南科技学院, 2014. RUAN T. Molecular epidemiological study of NDV in some areas of Henan and the application of NDV HN major antigenic region protein in antibody detection[D]. Xinxiang: Henan Institute of Science and Technology, 2014. |
| [37] |
郭威, 胡仕凤, 王卫国, 何世成, 唐小明, 林源, 鲁杏华, 王昌建. 环洞庭湖地区活禽市场鸭源NDV的分离鉴定及遗传演化分析[J]. 动物医学进展, 2015, 36(11): 69-73. DOI:10.16437/j.cnki.1007-5038.2015.11.015 GUO W, HU S F, WANG W G, HE S C, TANG X M, LIN Y, LU X H, WANG C J. Isolation and identification of duck-derived NDV in live poultry markets around Dongting Lake and genetic evolutionary analysis[J]. Advances in Animal Medicine, 2015, 36(11): 69-73. DOI:10.16437/j.cnki.1007-5038.2015.11.015 |
| [38] |
白晓. 9株不同来源的NDV遗传进化分析及其致病性研究[D]. 保定: 河北农业大学, 2019. BAI X. Genetic evolutionary analysis of nine NDV strains from different sources and their pathogenicity[D]. Baoding: Hebei Agricultural University, 2019. |
| [39] |
刘华雷, 郑东霞, 吕燕, 王静静, 赵云玲, 戈胜强, 王晓亮, 李知新, 张学军, 王福江, 门生宝, 王志亮. 宁夏地区鸡群新城疫流行率横断面调查[J]. 中国动物检疫, 2015, 32(10): 1-5. LIU H L, ZHENG D X, LYU Y, WANG J J, ZHAO Y L, GE S Q, WANG X L, LI Z X, ZHANG X J, WANG F J, MEN S B, WANG Z L. Cross-sectional survey on the prevalence of Newcastle disease in chickens in Ningxia region[J]. China Animal Quarantine, 2015, 32(10): 1-5. |
| [40] |
崔京腾. 鸭源新城疫病毒的分离鉴定及其对SPF鸭免疫机能的影响[D]. 泰安: 山东农业大学, 2013. CUI J T. Isolation and identification of duck-derived Newcastle disease virus and its effect on the immune function of SPF ducks[D]. Taian: Shandong Agricultural University, 2013. |
| [41] |
魏润宇. 2016-2017年我国部分地区NDV病原学监测及两种快速检测方法的建立[D]. 扬州: 扬州大学, 2018. WEI R Y. Pathogenetic surveillance of NDV and establishment of two rapid detection methods in some regions of China in 2016-2017[D]. Yangzhou: Yangzhou University, 2018. |
| [42] |
岳旭龙, 陈礼朋, 李双亮, 高文明, 张宇耕, 崔保安, 李新生. 2010-2011年河南省部分地区H9亚型禽流感、新城疫和鸡传染性支气管炎的流行病学调查[J]. 中国农学通报, 2012, 28(8): 72-76. YUE X L, CHEN L P, LI S L, GAO W M, ZHANG Y G, CUI B A, LI X S. Epidemiological survey of avian influenza H9 subtype, Newcastle disease and chicken infectious bronchitis in some areas of Henan Province in 2010-2011[J]. China Agronomy Bulletin, 2012, 28(8): 72-76. |
| [43] |
李剑静, 戴光文, 植枝林, 黄海英, 潘艳. 2019-2020年岑溪古典鸡禽流感和新城疫流行病学调查[J]. 湖北农业科学, 2021, 60(S1): 295-297. DOI:10.14088/j.cnki.issn0439-8114.2021.S1.079 LI J J, DAI G W, ZHI Z L, HUANG H Y, PAN Y. Epidemiological survey of avian influenza and Newcastle disease in classical chickens in Cenxi from 2019 to 2020[J]. Hubei Agricultural Sciences, 2021, 60(S1): 295-297. DOI:10.14088/j.cnki.issn0439-8114.2021.S1.079 |
| [44] |
廖飞, 杨先富, 赵孝木, 莫兴虎, 黄增荣. 黔东南小香鸡禽流感、新城疫流行病学调查[J]. 贵州畜牧兽医, 2018, 42(2): 44-45. LIAO F, YANG X F, ZHAO X M, MO X H, HUANG Z R. Epidemiological investigation of avian influenza and Newcastle disease in small aromatic chickens in Qiandongnan[J]. Guizhou Animal Husbandry and Veterinary Medicine, 2018, 42(2): 44-45. |
| [45] |
马弘财, 元振杰, 曾江勇. 西藏拉萨市H9亚型禽流感和新城疫流行病学调查与分析[J]. 现代畜牧兽医, 2019(12): 46-48. MA H C, YUAN Z J, ZENG J Y. Epidemiological investigation and analysis of avian influenza H9 subtype and Newcastle disease in Lhasa, Tibet[J]. Modern Animal Husbandry and Veterinary Medicine, 2019(12): 46-48. |
| [46] |
王倩, 刘华, 何长生, 朱良强, 占松鹤, 徐发荣, 沈朝建. 安徽省6个地市规模鸡场新城疫流行率估计及风险因素分析[J]. 中国动物检疫, 2021, 38(1): 24-29. WANG Q, LIU H, HE C S, ZHU L Q, ZHAN S H, XU F R, SHEN C J. Estimation of Newcastle disease prevalence and analysis of risk factors in large-scale chicken farms in six cities of Anhui Province[J]. China Animal Quarantine, 2021, 38(1): 24-29. |
| [47] |
温肖会, 杨彩娟, 翟少伦, 涂杜, 周秀蓉, 贾春玲, 吕殿红, 魏文康. 粤北地区H9N2亚型禽流感和新城疫流行情况调查[J]. 动物医学进展, 2019, 40(12): 140-144. DOI:10.16437/j.cnki.1007-5038.2019.12.030 WEN X H, YANG C J, ZHAI S L, TU D, ZHOU X R, JIA C L, LYU D H, WEI W K. Survey on the prevalence of H9N2 subtype of avian influenza and Newcastle disease in northern Guangdong[J]. Advances in Animal Medicine, 2019, 40(12): 140-144. DOI:10.16437/j.cnki.1007-5038.2019.12.030 |
| [48] |
朱钱龙, 张鸿, 刘平, 曾作财, 尧国荣. 江西省宜春地区鸡新城疫流行情况调查[J]. 畜牧与饲料科学, 2018, 39(8): 96-100. DOI:10.16003/j.cnki.issn1672-5190.2018.08.025 ZHU Q L, ZHANG H, LIU P, ZENG Z C, YAO G R. Survey on the prevalence of Newcastle disease in chickens in Yichun, Jiangxi Province[J]. Livestock and Feed Science, 2018, 39(8): 96-100. DOI:10.16003/j.cnki.issn1672-5190.2018.08.025 |
| [49] |
杨涛, 韦欣捷, 陈琼, 赵冉, 许淑娟. 厦门市规模禽场新城疫群流行率横断面研究估计[J]. 中国动物检疫, 2014, 31(11): 26-28, 32. YANG T, WEI X J, CHEN Q, ZHAO R, XU S J. A cross-sectional study to estimate the prevalence of Newcastle disease herd in large-scale poultry farms in Xiamen[J]. China Animal Quarantine, 2014, 31(11): 26-28, 32. |
| [50] |
刘浩. 陕西省鸡新城疫流行情况的病原学检测及其免疫效果分析[D]. 杨凌: 西北农林科技大学, 2014. LIU H. Pathogenetic detection of the prevalence of Newcastle disease in chickens in Shaanxi Province and its immunization effect analysis[D]. Yangling: Northwest Agriculture and Forestry University of Science and Technology, 2014. |
| [51] |
SHIRVANI E, SAMAL S K. Newcastle disease virus as a vaccine vector for SARS-CoV-2[J]. Pathogens, 2020, 9(8): 619. DOI:10.3390/pathogens9080619 |
| [52] |
O IE. Newcastle disease[J]. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals (Mammals, Birds and Bees), 2008(1): 576-589. |
| [53] |
姚舜禹, 张丽琳. 新城疫疫苗研究进展[J]. 生物技术进展, 2020, 10(5): 470-478. DOI:10.19586/j.2095-2341.2020.0052 YAO S Y, ZHANG L L. Advances in Newcastle disease vaccine research[J]. Advances in Biotechnology, 2020, 10(5): 470-478. DOI:10.19586/j.2095-2341.2020.0052 |
| [54] |
张婕, 李红娟. 大通县2013年禽流感、鸡新城疫流行病学调查报告[J]. 畜禽业, 2014(4): 70. DOI:10.19567/j.cnki.1008-0414.2014.04.047 ZHANG J, LI H J. Epidemiological investigation report of avian influenza and chicken Newcastle disease in Datong County in 2013[J]. Livestock and Poultry Industry, 2014(4): 70. DOI:10.19567/j.cnki.1008-0414.2014.04.047 |
| [55] |
袁仁强, 陈瑶生, 刘小红. 广东省生猪产业发展历史演变、问题与对策[J]. 广东农业科学, 2022, 49(5): 142-149. DOI:10.16768/j.issn.1004-874X.2022.05.017 YUAN R Q, CHEN Y S, LIU X H. Historical evolution, problems and countermeasures of pig industry development in Guangdong Province[J]. Guangdong Agricultural Sciences, 2022, 49(5): 142-149. DOI:10.16768/j.issn.1004-874X.2022.05.017 |
| [56] |
艾惠. F基因起始密码子突变对Class Ⅰ类NDV生物学特性的影响[D]. 北京: 中国农业科学院, 2020. DOI: 10.27630/d.cnki.gznky.2020.000758. AI H. Effect of mutation of F gene start codon on biological properties of Class Ⅰ NDV[D]. Beijing: Chinese Academy of Agricultural Sciences, 2020. DOI: 10.27630/d.cnki.gznky.2020.000758. |
| [57] |
MAMO T, YIMER L. Serological investigation of Newcastle disease in selected districts of Buno Bedelle Zone, Ethiopia[J]. Veterinary Medicine: Research & Reports, 2021, 12: 253-259. DOI:10.2147/VMRR.S327892 |
(责任编辑 崔建勋)