 2024, Vol. 51
2024, Vol. 51文章信息
引用本文 |

基金项目
- 广东省重点领域研发计划(2020B020215001);广东特支计划科技创新青年拔尖人才项目(2014TQ01N140)
作者简介
- 黄荣(1990—),男,博士,助理研究员,研究方向为林木遗传育种,E-mail:huangrong@sinogaf.cn.
通讯作者
- 郑会全(1982—),男,博士,教授级高级工程师,研究方向为林木遗传育种,E-mail:zhenghq@sinogaf.cn.
文章历史
- 收稿日期:2023-11-18




【研究意义】杉木〔Cunninghamia lanceolata (Lamb.) Hook.〕属柏科(Cupressaceae)杉木属(Cunninghamia)常绿乔木,是我国特色乡土针叶树种[1-2]。杉木不仅生长迅速、材质优良,且无明显病虫害,具有良好的经济和生态效益,已成为华南地区重要的商品用材与生态造林树种[2-3]。子代测定及评价是进行林木生长和材质性状评价和选择的重要方法[4]。与半同胞子代测定相比,全同胞子代测定能更精确地测定优质树种或无性系的遗传素质[5],可为多世代育种提供优良亲本,为良种选择提供可能。因此,开展杉木全同胞子代遗传测定和评价可有效选择高世代亲本杂交组合及优良育种材料,不断推进杉木高世代育种进程。【前人研究进展】自20世纪50年代开始,国内研究者对杉木进行系统研究,其中遗传改良领域一直备受重视。通过一系列科研攻关,杉木遗传育种工作在遗传资源收集、种源试验、种子园技术、优良家系选择、无性系选育等方面取得系列卓著的研究成果[2, 6-8]。杂交育种可以将双亲优良性状基因整合到杂交后代中,创造出符合生产需要的新基因型[9],是杉木遗传育种的重要途径。从20世纪70年代起,研究者认识到杉木种内杂种优势,并开展大量杉木杂交育种工作,使得杉木杂交育种得到了极大发展,促进了杉木遗传改良[6-7, 10-11]。【本研究切入点】亲本配合力是决定杂交后代优劣的重要标准之一,利用亲本配合力可筛选出遗传品质更为优异的杂交品系。随着杉木育种进程不断向高世代推进,本研究项目组从广东杉木第二代改良种子园亲本中筛选生长和结实表现好的核心亲本10个(cx38、cx455、cx539、cx569、cx570、cx571、cx573、cx574、cx576、cx580)[12],其中2个(cx569、cx570)入选第三代育种速生优质(生长和木材基本密度皆优)杉木亲本[13]。鉴于此,本研究利用这些核心亲本进行强化杂交,获得系列杂交全同胞家系,并进行多地点全同胞子代测定,研究其8年生的生长和木材基本密度遗传变异规律,筛选优异家系。【拟解决的关键问题】研究分析杉木全同胞家系的生长和木材基本密度在不同地点、不同家系间的遗传变异规律,以期选择生长材性皆优异的亲本杂交组合及优良育种材料。
1 材料与方法 1.1 试验地概况试验地共3处,分别位于广东北部和西北部,具体设在韶关市国有小坑林场(简称“小坑林场”)、韶关市国有九曲水林场(简称“九曲水林场”)、肇庆市国有大坑山林场(简称“大坑山林场”)。其中,小坑林场位于韶关市曲江区(24°41' N,113°51' E),属亚热带气候,年降雨量1 532 mm,全年平均气温20.2 ℃,土壤为红壤、肥力中等,试验地位于山坡中下部,为杉木林采伐迹地;九曲水林场位于韶关市翁源县(24°20' N,113°8' E),亚热带气候,年降雨量1 693 mm,全年平均气温20.6 ℃,土壤为黄壤、肥力中等,试验地位于山坡中部,为杉木林采伐迹地;大坑山林场位于肇庆市怀集县(23°45' N,112°11' E),亚热带气候,年降雨量1 693 mm,全年平均气温20.9 ℃,土壤为赤红壤、肥力中等,试验地位于山坡中下部,为杉木林采伐迹地。
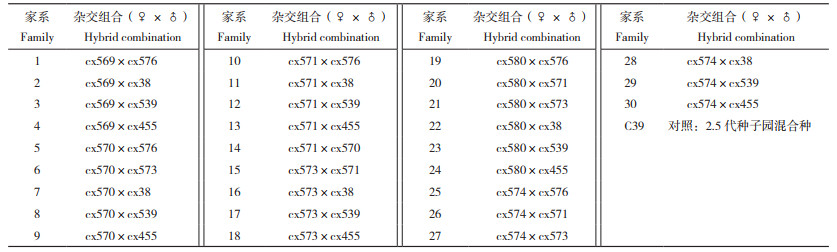
1.2 试验材料参试家系由广东杉木第二代改良种子园(小坑林场2.5代种子园)10个核心亲本,其中6个作为母本(种子园编号cx569、cx570、cx571、cx573、cx574、cx580)、7个作为父本(种子园编号cx38、cx455、cx539、cx570、cx571、cx573、cx576)定向交配创制而成,共计30个杂交组合(表 1),所获杂交组合种子翌年育成全同胞子代家系1年生裸根苗,同时以小坑林场杉木2.5代种子园混合种1年生裸根苗为对照,参照广东省地方标准DB44/T 778《杉木丰产林培育技术规程》,分别于小坑林场、九曲水林场、大坑山林场等地点营造试验林。试验采用随机完全区组设计,5个区组,2株小区,株行距2 m × 2 m。
|
1.3 性状测定
根据广东省地方标准DB44/T 1143《商品林经营管理规范》对速生丰产林、短周期工业原料用材林进行主要树种成熟龄的划分,杉木速生丰产林7~9年生属中龄林,已达半个轮伐期。在测定林8年生时(中龄林),分别对各试验点测定林的每木树高、胸径(DBH)进行测量。随后,根据公式V=0.00005877042 × H0.89646157 × D1.9699831,计算杉木单株材积[14]。同时,使用树木生长锥钻取每木胸径处(1.3 m高)木芯,采用饱和含水率法[15]测定其木材基本密度(WBD)。
1.4 数据分析首先采用R3.6.0平台并在R studio环境下对树高、胸径、材积和木材基本密度数据进行方差分析、表型变异系数、遗传变异系数和性状间相关系数分析等。参照林元震[16]的方法,采用亲本模型和个体模型分布开展数据统计分析。以单株观测值为基本数据,采用线性混合模型开展统计分析,数据分析工作在R3.6.0中的ASReml包进行方差分类估算、BLUP育种值预测[16]。在遗传力估算方面,计算家系遗传力采用公式 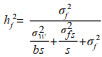
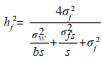
各性状现实增益计算采用公式:△(%)=(S/C)×100(S为选择差,C为对照测定值)。为筛选生长和木材基本密度表现皆优家系,分别以各家系(包括种子园品系)树高、胸径、单株材积、木材基本密度平均值为纵坐标,变异系数为横坐标绘制基因分组图,并以所有参试家系的性状总平均值与平均变异系数将图分为4组,凡某家系性状均值高于总平均值,而变异系数小于总平均变异系数,认为是具有理想基因型的稳定家系[17-18]。
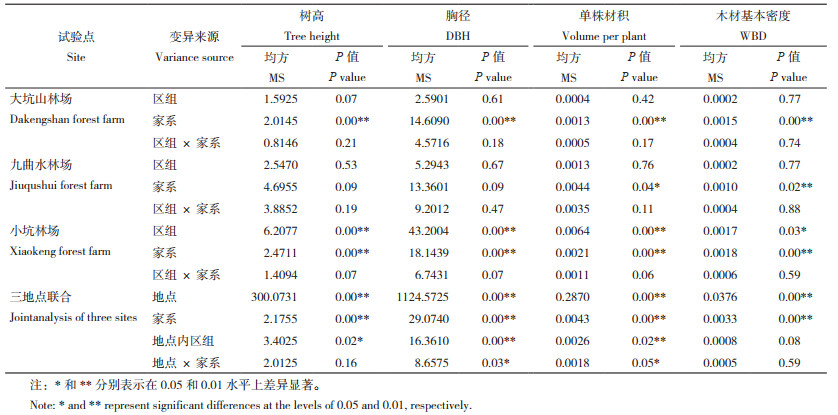
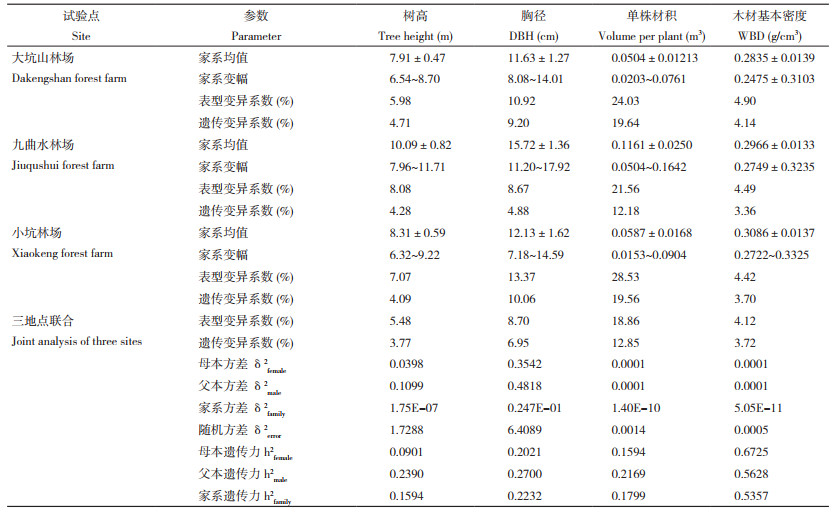
2 结果与分析 2.1 参试品系生长和木材基本密度的遗传变异对杉木30个全同胞子代家系在不同区域(大坑山林场、九曲水林场、小坑林场)8年生的生长材质表现进行系统测定,单地点方差分析结果(表 2)表明,在大坑山林场、九曲水林场、小坑林场3个试验点,单株材积和木材基本密度的家系效应差异均显著(P<0.05)或极显著(P<0.01),而大坑山林场和小坑林场的树高、胸径的家系效应也均达到极显著差异水平。三地点联合分析结果(表 2)也显示,树高、胸径、单株材积、木材基本密度等性状的家系效应差异达极显著(P<0.01)。以上结果表明,参试全同胞家系之间存在着真实的遗传差异。
|
对参试家系变异情况进行分析发现(表 3),测试点大坑山林场的树高、胸径、单株材积、木材基本密度性状变幅分别介于6.54~8.70 m、8.08~14.01 cm、0.0203~0.0761 m3、0.2475~0.3103 g/cm3间,九曲水林场各性状变化区间分别为7.96~11.71 m、11.20~17.92 cm、0.0504~ 0.1642 m3、0.2749~0.3235 g/cm3,而小坑林场树高、胸径、单株材积、木材基本密度性状则分别在6.32~9.22 m、7.18~14.59 cm、0.0153~0.0904 m3、0.2722~0.3325 g/cm3范围,由此可见,家系间生长(树高、胸径、单株材积)和木材基本密度表现变异广泛。尤其是各地点单株材积变异均较大,表型变异系数和遗传变异系数分别达到了21.56%~28.53%、12.18%~19.64%;其次为胸径、树高、木材基本密度的变异。此外,综合3处测试点生长和木材基本密度表现,发现树高、胸径、单株材积、木材基本密度跨区域表型变异系数为5.48%、8.70%、18.86%、4.12%,遗传变异系数则达3.77%、6.95%、12.85%、3.72%。在遗传力方面,杉木生长性状家系遗传力介于0.1594~0.2232间,而母本遗传力、父本遗传力则分别介于0.0901~0.2021、0.2169~0.2700间;对于木材基本密度而言,无论是家系遗传力还是母本和父本遗传力均属较高水平,分别为0.5357、0.6725、0.5628。
|
2.2 生长和木材基本密度间的相关性
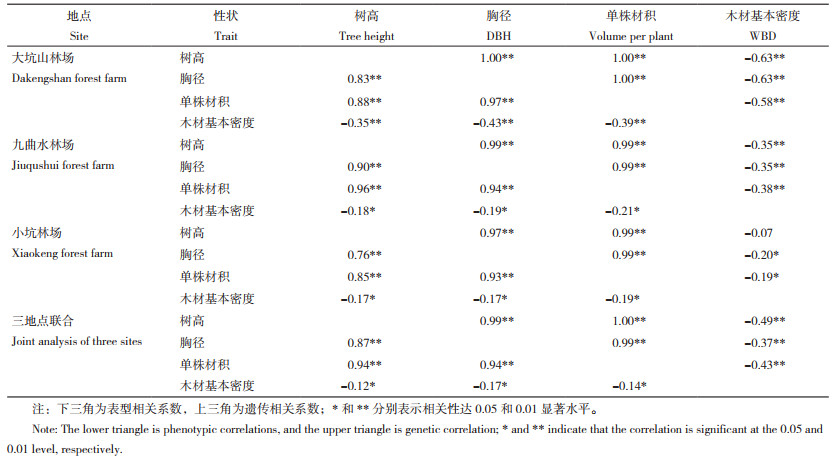
对测试群体不同生长(树高、胸径、单株材积)、木材基本密度性状单地点和多地点联合的表型相关系数和遗传相关系数进行计算发现(表 4),树高、胸径和单株材积3个生长性状无论在表型还是在遗传层次上均存在极显著的正相关关联,相关系数均在0.75以上。然而,生长性状与木材基本密度间无论从表型还是遗传方面均呈现出负相关关系,且达到显著(在表型相关方面)或极显著(在遗传相关方面)水平。
|
2.3 参试杂交组合(全同胞家系)亲本育种值及其特殊配合力估算
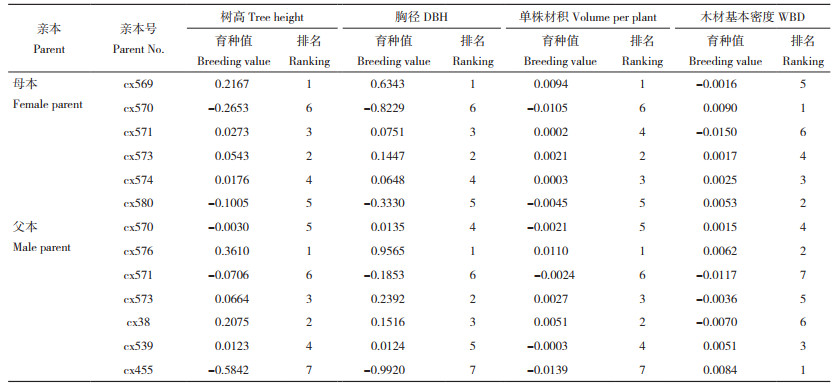
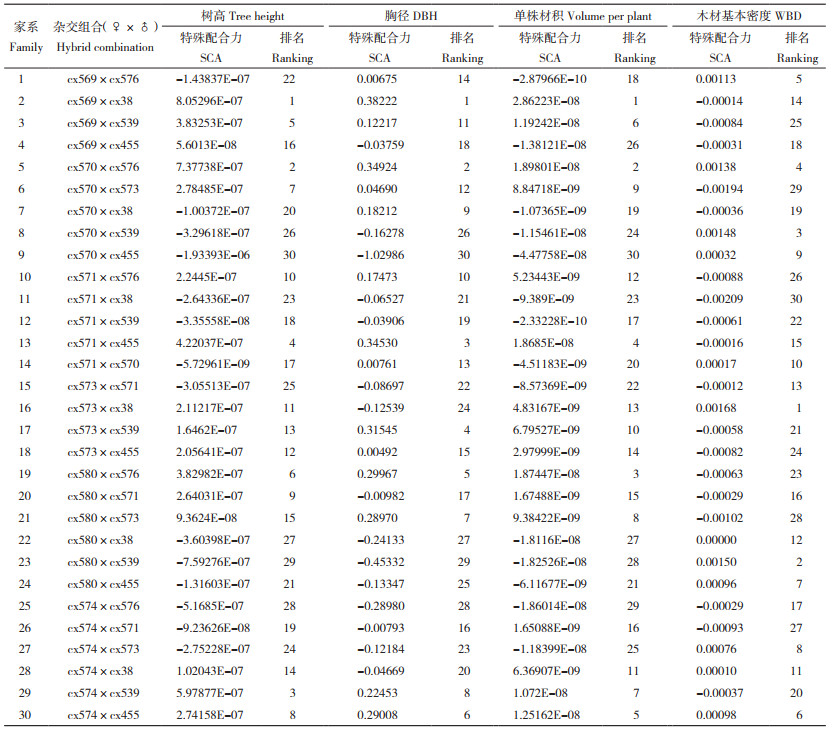
基于全同胞家系的生长性状和木材基本密度测定值估算全同胞家系母本和父本在树高、胸径、单株材积和木材基本密度等性状方面的育种值。结果(表 5)显示,母本间、父本间育种值存在较大差异,其中母本cx569、cx573、cx571树高育种值排名靠前(前3位),母本cx569、cx573在胸径和单株材积方面的育种值均排名第1和第2,木材基本密度育种值排名前三位的母本分别为cx570、cx580和cx574。而父本上,cx576、cx38、cx573生长性状育种值位列前茅(前3位),cx455、cx576、cx539的木材基本密度育种值更高(前3位)。此外,按亲本模型估算各杂交组合的特殊配合力发现,参试杂交组合树高、胸径、单株材积、木材基本密度性状的特殊配合力分别介于-1.93393E-06~8.05296E-07、-1.029863034~0.382218308、-4.47758E-08~2.86223E-08、-0.00209~0.00168之间(表 6),存在较大的差异。在树高、胸径和单株材积方面具有较高特殊配合力的杂交组合其木材基本密度特殊配合力往往较低,但也有例外,如杂交组合cx570×cx576和cx574×cx455,这2个组合在生长、木材基本密度性状方面均具有较高的特殊配合力。
|
|
2.4 优良杂交组合(全同胞家系)选择及其现实增益
将不同家系在3个测试地点的平均树高、胸径、单株材积作为生长性状考察指标,变异系数为稳定性指标,同时以群体均值为阈值,对参试的30个全同胞子代家系进行分组(图 1)。显然,就树高性状而言,表现较优且稳定的家系有2、5、10、14、17、19、21、25、29等;就胸径性状而言,2、5、10、14、17、19、21、25、27、29等家系表现稳定且突出;将单株材积作为生长性状综合考察指标,2、5、10、17、19、21、25、29家系具有丰产、稳产特性。在材质选择方面,图 1D划分出了木材基本密度高且具有较好稳定性的家系(4、5、9、16、18、23、24、27、29)。对参试家系生长材质作联合选择,家系5(杂交组合cx570×cx576)和家系29(杂交组合cx574×cx539)可初步入选。

|
| 虚线表征群体均值 Dotted line represents mean value 图 1 参试杉木全同胞家系生长和材质分组 Fig. 1 Grouping accding to growth and wood property traits of Cunninghamia lanceolata full-sib families |
较群体均值,家系5、家系29在3个测定地点的单株材积增益分别可达11.66%~22.37%(平均15.82%)、-9.36%~24.60%(平均3.78%),而木材基本密度增益则分别为5.92%~9.18%(平均7.25%)、0.38%~1.78%(平均1.35%),树高和胸径增益分别为4.52%~7.24%(平均6.02%)、-1.37%~8.17%(平均4.04%)和5.89%~11.65%(平均8.21%)、-2.89%~9.76%(平均5.06%)。鉴于系列全同胞家系测试时还增加了2.5代种子园混合种为对照,故以之为标准,进一步发现家系5、家系29在3个测定地点较2.5代种子园混合种的单株材积增益分别可达23.49%~66.43%(平均34.56%)、0.25%~65.43%(平均20.57%),木材基本密度增益分别为-1.02%~11.79%(平均5.35%)、-5.04%~4.24%(-0.45%)、树高增益分别达10.24%~35.63%(平均26.47%)、11.20%~36.69%(平均23.93%)、胸径增益则分别可达13.17%~15.62%(平均14.25%)、6.03%~ 16.80%(平均10.87%)。
3 讨论本研究对杉木10个核心亲本杂交得到的30个全同胞家系8年生子代开展了遗传测定和分析,发现参试全同胞家系的生长(树高、胸径、单株材积)和材质(木材基本密度)性状均达显著(P<0.05)或极显著水平(P<0.01),并且这些性状尤其生长性状的变异系数较大,由此说明,生长和木材基本密度在家系间存在广泛的遗传差异,为下一步杉木优良家系的选择奠定了材料基础。表型变异受到自身遗传和环境因素共同影响,可通过遗传参数估算解析遗传和环境对其贡献大小,其中遗传力可以反映性状遗传给子代的几率,即遗传力越大,性状稳定遗传的几率越高[6]。本研究参试全同胞家系的生长性状家系遗传力为中低遗传力,而木材基本密度家系遗传力为中等遗传力,表明杉木全同胞家系生长尤其木材基本密度受一定程度遗传控制,特别木材基本密度具有更为稳定的遗传能力,因此这些生长和木材基本密度可作为家系选择的主要性状[4, 7, 19]。林木性状间相关性研究揭示性状之间的相关程度,从而为林木遗传改良提供重要参考[20]。本研究中,杉木生长性状间呈现极显著正相关,而生长性状和木材基本密度为显著或极显著负相关,与以往诸多研究结果一致,表明杉木的生长与木材基本密度可能相互独立遗传[20-22]。这意味着在杉木育种过程中强化生长性状的改良将会导致木材基本密度下降,因此,在杉木改良工作中应该同时兼顾生长和木材基本密度[21-23]。
本研究综合生长和材质对30个参试家系进行选择,筛选出家系5和29,其平均树高、胸径、单株材积、基本密度分别达8.95~9.17 m、13.51~14.03 cm、0.0764~0.0832 m3、0.3000~0.3173 g/cm3。育种值是家系性状综合表现的重要参数,可真实反映家系遗传品质优劣,提高选择的精确性[7]。入选的2个家系亲本育种值排名较靠前,尤其家系5的父本生长性状育种值排名第1,木材基本密度的育种值排名第2,其母本木材基本密度排名第1,表明它们亲本遗传品质优异。同时,这两个家系的亲本在生长、木材基本密度性状方面均具有较高的特殊配合力,尤其是杂交组合cx570×cx576(即家系5亲本组合),表明这两对亲本组合较大概率可以产出生长和材质皆优的后代。遗传增益可以反映育成群体相较现有群体目的性状提高的程度,是评价育种效果的重要参数[21]。与对照2.5代种子园混合种相比,家系5和家系29在3个测定地点的单株材积平均增益分别达34.56%和20.57%,木材基本密度平均增益则分别为5.35% 和-0.45%。综合以上,家系5和29两者中,家系5具有更高的生长和木材基本密度增益,是更理想的速生优质品系(家系)。
4 结论对杉木全同胞家系8年生生长和木材基本密度遗传变异规律进行研究,并对其育种值和特殊配合力进行估算。研究结果表明杉木全同胞家系间生长和木材基本密度差异明显,且遗传力较高,可作为家系选择的主要性状。综合育种值排名、杂交组合的特殊配合力以及生长性状和木材基本密度表现,最终筛选出2个速生优质型家系(家系5和29)。尤其家系5,相比群体均值树高、胸径、单株材积及木材基本密度增益平均为6.02%、8.21%、15.82%、7.25%,而相比对照(2.5代种子园混合种)增益平均为26.47%、14.25%、34.56%、5.35%,是更理想的速生优质家系。
| [1] |
YANG Y, FERGUSON D K, LIU B, MAO K S, GAO L M, ZHANG S Z, WAN T, RUSHFORTH K, ZHANG Z X. Recent advances on phylogenomics of gymnosperms and an updated classification[J]. Plant Diversity, 2022, 44(4): 340-350. DOI:10.1016/j.pld.2022.05 |
| [2] |
王润辉, 胡德活, 韦如萍, 晏姝, 郑会全. 杉木育种亲本生长和结实性状早期测评与选择[J]. 热带亚热带植物学报, 2022, 30(2): 195-201. DOI:10.11926/jtsb.4457 WANG R H, HU D H, WEI R P, YAN S, ZHENG H Q. Early evaluation and selection of the Chinese fir breeding parents based on growth and cone production traits[J]. Journal of Tropical and Subtropical Botany, 2022, 30(2): 195-201. DOI:10.11926/jtsb.4457 |
| [3] |
邓厚银, 晏姝, 王润辉, 胡德活, 郑会全, 韦如萍, 梁机. 粤北不同林龄杉木人工林下土壤层及枯落物层持水能力研究[J]. 广东农业科学, 2019, 46(2): 51-58. DOI:10.16768/j.issn.1004-874X.2019.02.008 DENG H Y, YAN S, WANG R H, HU D H, ZHENG H Q, WEI R P, LIANG J. Study on water-holding capacity of soil layer and litter layer of Chinese fir plantations with different stand ages in northern Guangdong[J]. Guangdong Agricultural Sciences, 2019, 46(2): 51-58. DOI:10.16768/j.issn.1004-874X.2019.02.008 |
| [4] |
晏姝, 胡德活, 郑会全, 王润辉, 韦如萍, 林军. 16年生杉木2代种子园家系区域测定分析[J]. 森林与环境学报, 2018, 38(4): 414-418. DOI:10.13324/j.cnki.jfcf.2018.04.005 YAN S, HU D H, ZHENG H Q, WANG R H, WEI R P, LIN J. Regional experiment on 16-year-old families of the 2nd generation seed orchard of Cunninghamia lanceolata[J]. Journal of Forest and Environment, 2018, 38(4): 414-418. DOI:10.13324/j.cnki.jfcf.2018.04.005 |
| [5] |
ASTLES P A, MOORE A J, PREZIOSI R F. A comparison of methods to estimate cross-environment genetic correlations[J]. Journal of Evolutionary, 2006, 19(1): 114-122. DOI:10.1111/j.1420-9101.2005.00997.x |
| [6] |
陈代喜, 陈晓明, 蓝肖, 戴俊, 罗启亮, 莫宗恒, 肖建军. 杉木全同胞子代遗传测定与优良种质选择[J]. 广西林业科学, 2016, 45(4): 347-351. DOI:10.19692/j.cnki.gfs.2016.04.001 CHEN D X, CHEN X M, LAN X, DAI J, LUO Q L, MO Z H, XIAO J J. Full-sib progeny genetic test and selection of superior germplasm of Cunninghamia lanceolata[J]. Guangxi Forestry Science, 2016, 45(4): 347-351. DOI:10.19692/j.cnki.gfs.2016.04.001 |
| [7] |
段爱国, 孙建军, 邓宗富, 张建国. 杉木第三代亲本杂交子代生长遗传变异及选择研究[J]. 江西农业大学学报, 2022, 44(2): 261-270. DOI:10.13836/j.jjau.2022027 DUAN A G, SUN J J, DENG Z F, ZHANG J G. Studies on genetic variation and evaluation of hybrid offspring of the parents in the third generation seed orchards of Chinese fir[J]. Acta Agriculturae Universitatis Jiangxiensis, 2022, 44(2): 261-270. DOI:10.13836/j.jjau.2022027 |
| [8] |
HUANG R, WANG R, WEI R, YAN S, WU G, ZHENG H. Selection for both growth and wood properties in Chinese fir breeding parents based on a 6-year grafted clone test[J]. Forests, 2023, 14: 1774. DOI:10.3390/f14091774 |
| [9] |
WANG S, ZHAO S, HUANG Z, YIN L, HU J, LI J, LIU Y, RONG C. Development of a highly productive strain of Pleurotus tuoliensis for commercial cultivation by crossbreeding[J]. Scientia Horticulturae, 2018, 234: 110-115. DOI:10.1016/j.scienta.2018.02.001 |
| [10] |
黄越茂, 王润辉, 郑会全, 胡德活, 植毓永, 蔡维就. 杉木杂交子代遗传测定分析[J]. 现代农业科技, 2017(4): 125-126. DOI:10.3969/j.issn.1007-5739.2017.04.077 HUANG Y M, WANG R H, ZHENG H Q, HU D H, ZHI Y Y, CAI W J. Analysis on genetic test of hybrid progeny of Cunninghamia lanceolata(Lamb.) Hook.[J]. Modern Agricultural Science and Technology, 2017(4): 125-126. DOI:10.3969/j.issn.1007-5739.2017.04.077 |
| [11] |
邓厚银, 胡德活, 林军, 赖旭恩, 邓文剑, 韦如萍, 王润辉, 晏姝, 郑会全. 杉木半同胞子代胸径变异和大径材家系选择[J]. 热带亚热带植物学报, 2020, 28(5): 513-519. DOI:10.11926/jtsb.4205 DENG H Y, HU D H, LIN J, LAI X E, DENG W J, WEI R P, WANG R H, YAN S, ZHENG H Q. Diameter variation of Chinese fir half-sib progenies and selection for large-size timber families[J]. Journal of Tropical and Subtropical Botany, 2020, 28(5): 513-519. DOI:10.11926/jtsb.4205 |
| [12] |
胡德活, 林军, 王润辉, 郑会全, 梁瑞友, 晏姝, 韦如萍, 刘伟新, 赖旭恩, 吕宇宙, 李彭生. 杉木第二代改良种子园营建技术及应用成效[J]. 林业与环境科学, 2019, 35(1): 23-28. DOI:10.3969/j.issn.1006-4427.2019.01.005 HU D H, LIN J, WANG R H, ZHENG H Q, LIANG R Y, YAN S, WEI R P, LIU W X, LAI X E, LYU Y Z, LI P S. Establishment techniques for improved second generation seed orchard of Chinese f ir and its applying achievement[J]. Forestry and Environmental Science, 2019, 35(1): 23-28. DOI:10.3969/j.issn.1006-4427.2019.01.005 |
| [13] |
黄荣, 胡德活, 邓厚银, 王润辉, 韦如萍, 晏姝, 郑会全. 利用SNP标记解析速生优质杉木群体的遗传多样性与结构[J/OL]. 分子植物育种, 1-10[2023-11-18]. http://kns.cnki.net/kcms/detail/46.1068.S.20211130.1703.012.html. HUANG R, HU D H, DENG H Y, WANG R H, WEI R P, YAN S, ZHENG H Q. SNPs-based assessment of genetic diversity and genetic structure in elite Chinese fir[J/OL]. Molecular Plant Breeding, 1-10 [2023-11-18]. http://kns.cnki.net/kcms/detail/46.1068.S.20211130.1703.012.html. |
| [14] |
胡德活, 郝玉宝, 梁机, 郑会全, 王润辉, 韦如萍, 晏姝, 赖旭恩, 林军, 梁庆. 广东乐昌杉木种质资源库无性系生长与材质性状变异分析[J]. 西南林业大学学报, 2011, 31(6): 1-5. DOI:10.3969/j.issn.2095-1914.2011.06.001 HU D H, HAO Y B, LIANG J, ZHENG H Q, WANG R H, WEI R P, YAN S, LAI X E, LIN J, LIANG Q. Variation analysis on growth and wood quality traits of Cunninghamia lanceolata clones in Lechang germplasm bank[J]. Journal of Southwest Forestry University, 2011, 31(6): 1-5. DOI:10.3969/j.issn.2095-1914.2011.06.001 |
| [15] |
成俊卿. 木材学[M]. 北京: 中国林业出版社, 1985. CHENG J Q. Wood science[M]. Beijing: China Forestry Publishing House, 1985. |
| [16] |
林元震. R与ASReml-R统计学[M]. 北京: 中国林业出版社, 2016. LIN Y Z. R and ASReml-R statistics[M]. Beijing: China Forestry Publishing House, 2016. |
| [17] |
王润辉, 张伟红, 郑会全, 晏姝, 韦如萍, 胡德活, 植毓永, 罗平峰, 黄小平, 吴雄光. 乐昌含笑多点种源试验与优良种源选择[J]. 中南林业科技大学学报, 2015, 35(5): 16-21, 34. DOI:10.14067/j.cnki.1673-923x.2015.05.003 WANG R H, ZHANG W H, ZHENG H Q, YAN S, WEI R P, HU D H, ZHI Y Y, LUO P F, HUANG X P, WU X G. Study on mutil-site provenance trails and superior provenances selection of Michelia chapensis[J]. Journal of Central South University of Forestry & Technology, 2015, 35(5): 16-21, 34. DOI:10.14067/j.cnki.1673-923x.2015.05.003 |
| [18] |
张伟红, 王润辉, 郑会全, 晏姝, 韦如萍, 胡德活, 植毓永, 罗平峰, 黄小平, 吴雄光, 何汉波. 乐昌含笑优树多点子代测定及优良家系选择[J]. 浙江农林大学学报, 2015, 32(5): 763-769. DOI:10.11833/j.issn.2095-0756.2015.05.016 ZHANG W H, WANG R H, ZHENG H Q, YAN S, WEI R P, HU D H, ZHI Y Y, LUO P F, HUANG X P, WU X G, HE H B. Mutil-site family trails and selection of superior families for Michelia chapensis[J]. Journal of Zhejiang A & F University, 2015, 32(5): 763-769. DOI:10.11833/j.issn.2095-0756.2015.05.016 |
| [19] |
叶代全. 杉木第4代育种候选群体的12年生全同胞子代测定表现与选择[J]. 南京林业大学学报(自然科学版), 2022, 46(6): 240-250. DOI:10.12302/j.issn.1000-2006.202206050 YE D Q. Performances and selections on a 12-year-old full-sib progeny testing from one of the candidate population for the 4th generation Chinese fir breeding[J]. Journal of Nanjing Forestry University(Natural Sciences Edition), 2022, 46(6): 240-250. DOI:10.12302/j.issn.1000-2006.202206050 |
| [20] |
胡德活, 阮梓材, 钱志能, 黄越茂, 黄小平, 陈忠林. 杉木无性系木材密度遗传变异及其与生长性状的相关性[J]. 中南林业科技大学学报, 2004, 24(5): 24-27. DOI:10.3969/j.issn.1673-923x.2004.05.006 HU D H, RUAN Z C, QIAN Z N, HUANG Y M, HUANG X P, CHEN Z L. The genetics and variation of wood density in Chinese fir clones and its interaction to the growth characters[J]. Journal of Central South University of Forestry & Technology, 2004, 24(5): 24-27. DOI:10.3969/j.issn.1673-923x.2004.05.006 |
| [21] |
何贵平, 骆文坚, 金其祥, 冯建国, 冯建民, 洪长胜. 杉木无性系主要生长、材质性状遗传差异及无性系选择[J]. 江西农业大学学报, 2009, 31(1): 91-93. DOI:10.3969/j.issn.1000-2286.2009.01.020 HE G P, LUO W J, JIN Q X, FENG J G, FENG J M, HONG C S. Genetic difference and clone selection of main traits of growth and wood quality of Chinese fir clone[J]. Acta Agriculturae Universitatis Jiangxiensis, 2009, 31(1): 91-93. DOI:10.3969/j.issn.1000-2286.2009.01.020 |
| [22] |
赵林峰, 高建亮, 彭劲松, 姜小文. 3种类型杉木林分生长、密度和材质的综合评价[J]. 安徽农业大学学报, 2023, 50(1): 29-35. DOI:10.13610/j.cnki.1672-352x.20230315.021 ZHAO L F, GAO J L, PENG J S, JIANG X W. Comprehensive evaluation of the growth, density and texture of three types of Cunninghamia lanceolata[J]. Journal of Anhui Agricultural University, 2023, 50(1): 29-35. DOI:10.13610/j.cnki.1672-352x.20230315.021 |
| [23] |
黄金华. 秃杉中龄人工林凋落物量8年动态特征[J]. 广东农业科学, 2017, 44(9): 56-62. DOI:10.16768/j.issn.1004-874X.2017.09.009 HUANG J H. Dynamics of litter production in a middle-aged Taiwania cryptomerioides plantation over an 8-year period[J]. Guangdong Agricultural Sciences, 2017, 44(9): 56-62. DOI:10.16768/j.issn.1004-874X.2017.09.009 |
(责任编辑 马春敏)