 2022, Vol. 49
2022, Vol. 49文章信息
引用本文 |

基金项目
- 广东省科技计划项目(2022B0202060002);国家现代农业产业技术体系建设专项资金(CARS-Rice);广东省农业科学院农业优势产业学科团队建设项目(202101TD);广东省学科类重点实验室运行经费项目(2020B1212060047)
作者简介
-
王丰,博士、二级研究员,广东省农业科学院水稻研究所首席科学家、农业农村部华南优质稻遗传育种重点实验室主任。主持863计划、国家重点研发计划、广东省重大科技专项、广东省重点研发计划等项目20余项。主要从事杂交稻遗传与育种研究,育成杂交稻不育系泰丰A、荣丰A、吉丰A等22个通过鉴定;育成优质高产抗病杂交稻新品种70多个和广东丝苗米新品种1个通过审定;研究成果为国内10多家水稻种子企业发展提供了重要技术支撑,累计推广应用面积超过2亿亩。获各级科技奖励成果19项,其中国家科技进步特等奖和一等奖各1项;省级科学技术一等奖6项;获发明专利4项、植物新品种权18项。在国内外学术刊物上发表论文100多篇。
王丰(1963—),男,博士,研究员,研究方向为杂交水稻遗传育种,E-mail:fwang1631@163.com.
文章历史
- 收稿日期:2022-07-19



水稻是重要的粮食作物,对我国的粮食安全具有举足轻重的影响。20世纪70年代我国率先实现籼型杂交水稻三系法配套[1]。此后,水稻杂种优势利用技术在我国持续快速发展,杂交稻品种在南方稻区广泛推广与应用,种植面积不断扩大。南方稻区种植杂交稻的面积最高时占我国水稻种植总面积的80% 以上,为我国粮食增产增收和粮食安全做出了巨大贡献,取得了举世瞩目的成就[2]。
广东等华南稻区晚季仍具有光照强、日照短、温度高等特殊的光温生态条件,生产上客观需要种植弱感光型杂交稻品种[3-6]。目前,应用的弱感光型杂交稻品种主要有两大类,一类是由不育系含互作型弱感光基因(如博A、秋A、振丰A等)组配育成的弱感光型组合,另一类是由恢复系含互作型弱感光基因(如广恢3550、广恢55等)组配育成的弱感光型组合[6]。这些携带互作型弱感光基因的不育系或恢复系已应用多年,其品质、抗性等已不能完全适合当前生产和市场的需求,尤其在弱感光型迟熟杂交稻的主栽地华南沿海地区,台风雨多,白叶枯病发生重,严重影响其高产稳产。因此,亟需选育含互作型弱感光基因和抗白叶枯病的新不育系、恢复系和新组合。
杂交稻不育系和恢复系的选育是利用水稻杂种优势的关键环节。不育系选育所需时间较长[7],恢复系选育具有周期短、成效快的特点。同一恢复系与不同不育系通常能组配出不同熟期特色的杂交稻新组合,可满足不同稻作生态区域的生产需求。因此,恢复系的选育是提高杂交稻品质、产量、抗性和改变品种熟期的重要途径。本研究以弱感光型恢复系选育为重点,利用常规杂交和目标基因分子标记辅助选择技术,聚合互作感光基因和白叶枯病抗性基因,培育出抗白叶枯病且含弱感光互作基因的“R3550类型”[6]恢复系广恢1002,进而与不育系组配选育高产、优质、抗病的弱感光型杂交稻新组合,以促进华南稻区弱感光型杂交稻品种的更新换代,推进水稻杂种优势利用技术不断创新发展。
1 抗白叶枯病弱感光型杂交稻恢复系广恢1002的选育 1.1 广恢1002的选育过程2005年早季利用携带显性白叶枯病抗性基因Xa7的恢复系广恢806为母本,与广东省农业科学院水稻研究所育成的携带互作感光基因的“R3550类型”恢复系广恢55(来源于广恢3550// 粤龙占/ 广恢122)杂交;同年晚造栽种F1代并再与广恢55回交。2006年早季种BC1F1,晚季种F2代1 600株,成熟期筛选到综合性状优良(特别是叶厚直且后期耐寒性较强)的迟熟单株共9株。2007年早、晚季种植F3、F4株系,成熟期进行系谱选择;F4、F5代进行白叶枯病田间接种抗性鉴定,在稻瘟病区进行稻瘟病自然诱发抗性鉴定,选择抗白叶枯病、抗稻瘟病且迟熟的优良单株;F6、F7代继续进行白叶枯病田间接种抗性鉴定,以及稻瘟病自然诱发抗性鉴定,所有亲本中均携带LP1,且只利用分子标记(RM25和FB2)检测互作感光特性基因(LP2或LH2)[8-9],最后选择抗白叶枯病、抗稻瘟病和具有互作感光基因的优良单株与不育系吉丰A、安丰A、泰丰A等进行测恢测优及杂种F1代白叶枯病鉴定,对新选育出的苗头恢复系进行稻米外观与蒸煮食味品质鉴定。2009年(F8代)初步育成了抗白叶枯病含弱感光互作基因的迟熟新恢复系,2011年利用抗白叶枯病基因Xa7紧密连锁的SSR标记M3进行检测分析[8],选择Xa7为纯合基因型且同时携带纯合稳定互作感光特性基因的高配合力单株,定名为广恢1002。广恢1002的系谱如图 1所示。

|
| 图 1 广恢1002的系谱 Fig. 1 Pedigree of restorer R1002 |
1.2 广恢1002的特征特性
广恢1002具有互作感光特性,与多数三系不育系组配,其杂种F1均表现为弱感光迟熟特性。广恢1002在广州早造3月上旬播种,6月中旬抽穗,全生育期146 d左右;在广州晚造7月下旬播种,10月上旬抽穗,全生育期为115~117 d。广恢1002的叶片较为窄直,分蘖力强,有效穗较多,株叶形态好,制种花粉量大,易获高产。广恢1002品质较好,千粒质量22.7 g左右,米粒长6.3 mm,粒宽2.1 mm,米粒长宽比达到3.0,心腹白较少,直链淀粉含量较低。广恢1002配合力强,恢复谱广,与不同细胞质源的三系不育系组配,其杂种F1的结实率均很高,显示出很强的恢复力。在抗病性方面,由于广恢1002导入了抗白叶枯病基因Xa7,因此,其白叶枯病抗性较好,经接种白叶枯病Ⅳ型、Ⅸ型菌,抗性分别达到1级和3级。此外,广恢1002对细条病、稻瘟病和黑条矮缩病等均具有较好的田间抗性。
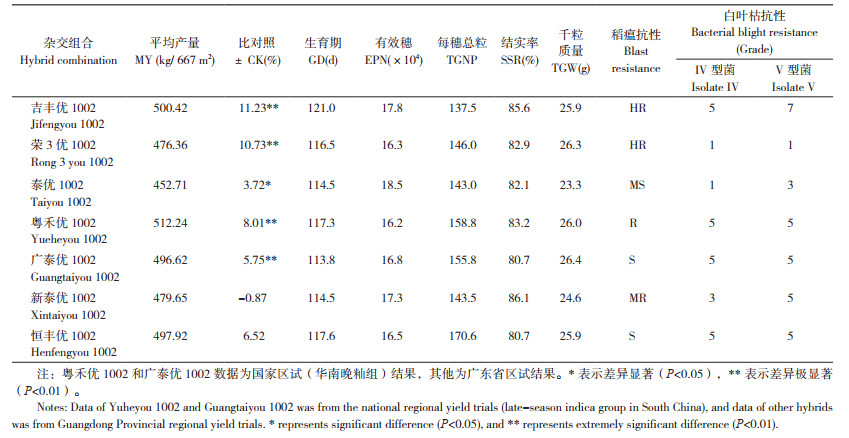
2 广恢1002系列杂交稻的特征特性 2.1 产量与抗性利用抗白叶枯病的恢复系广恢1002与不育系组配,先后育成了吉丰优1002、泰优1002、粤禾优1002、荣3优1002、广泰优1002、恒丰优1002和新泰优1002等7个组合,均为弱感光型晚籼迟熟杂交稻,已于2013、2017、2019、2020、2021年分别通过了广东或(和)国家品种审定。7个组合中,除新泰优1002外,其他6个组合在区试均表现出比对照增产达显著或极显著水平,显示出广恢1002很好的配合力与杂种优势。其中,区试产量最高的是粤禾优1002,两年平均每667 m2产量高达512.24 kg;但比对照增产幅度最大的是吉丰优1002,比对照博优998和博Ⅲ优273两年平均增产11.23%,增产达极显著水平。生育期最长的是吉丰优1002,最短的是广泰优1002,仅为113.8 d,与泰优1002和新泰优1002接近。在抗病性方面,荣3优1022的抗病性最好,不仅高抗稻瘟病,还抗白叶枯病。其他6个组合尽管人工接种鉴评结果对白叶枯病均表现为中感或感,但在大田生产上表现出很好的抗白叶枯病(图 2、图 3)。各组合的具体特征特性如表 1所示。

|
| 图 2 泰优1002大田抗白叶枯病表现 Fig. 2 Performance of Taiyou 1002 in resistance to bacterial blight in the fields |

|
| 图 3 吉丰优1002大田抗白叶枯病表现 Fig. 3 Performance of Jifeng you 1002 in resistance to bacterial blight in the fields |
|
2.2 稻米品质
广恢1002稻米品质较好,综合品质指标达到国标优质3级。利用其组配的杂交稻,除吉丰优1002和荣3优1002外,其他5个品种的稻米均达到国标优质3级或部标优质2级标准(表 2)。其中,泰优1002品质最好,不仅达到部优2级标准,而且在第三届全国优质稻品种食味鉴评中获得籼稻组金奖。广泰优1002的母本广泰A是一个优质的香型不育系,因此,广泰优1002的米饭不仅适口性与食味较好,而且带有清香味。

3 广恢1002系列杂交稻的综合评述 3.1 吉丰优1002
该组合是利用分子标记辅助选择技术将广谱抗瘟基因Pi1导入荣丰B后育成的三系不育系吉丰A与广恢1002组配选育而成的,2013年通过广东省品种审定(粤审稻2013040)。目前已引种至广西和海南等华南的西南部地区进行大面积推广应用。尽管米质未达优质等级,但非常符合粤、桂、琼等沿海地区人们的饮食习惯。该组合丰产性突出,抗倒性强,高抗稻瘟病,大田表现抗白叶枯病和细条病(图 3),在粤、桂、琼等台风雨频繁的沿海地区表现高产稳产,被农民称之为“无忧稻”。从2017年开始,吉丰优1002连续5年成为广东单品单造销量最大的弱感光型迟熟杂交稻组合,已累计推广超过20万hm2。2017年被认定为超级稻,2021—2022年被列为广东省农业主导品种,也是国家区试华南晚籼感光迟熟组的区试对照品种。
3.2 泰优1002该组合是利用优质不育系泰丰A与广恢1002组配育成的弱感光型迟熟三系优质杂交稻品种。2015、2016年晚造参加广东省区试和生产试验,于2017年通过广东省品种审定(粤审稻20170081)。泰优1002生育期较短,丰产性较好,对白叶枯病和细条病的田间抗性好(图 2),且稻米品质优,达到国标优质3级。2020年在国家第三届优质稻食味鉴评中获籼稻组金奖。2021— 2022年被列为广东省农业主导品种。在广东、广西晚造深受欢迎,已累计推广超过6万hm2。
3.3 粤禾优1002该组合利用携带抗稻瘟病主效基因Pi1的三系不育系粤禾A与广恢1002组配而成的,2019年通过国家品种审定(国审稻20190156)和广东省品种审定(粤审稻20190032)。2017、2018年参加华南感光晚籼组水稻联合体区域试验,两年区域试验平均产量7.68 t/hm2,比对照博优998增产8.01%;生产试验平均产量8.10 t/hm2,比对照博优998增产9.95%。2017、2018年同时推荐参加广东省晚造区试,平均产量分别为7.02、7.17 t/hm2,比对照广8优169分别增产10.02% 和8.12%,增产均达极显著水平,增产点比例分别为100%、90.91%。2018年晚造参加广东省生产试验,平均产量7.01 t/hm2,比广8优169增产5.92%。稻瘟病全群抗性频率为75.0%~89.66%,对中B群、中C群的抗性频率分别为75.00%~90.48% 和100%,病圃鉴定叶瘟1.0~2.0级、穗瘟1.7~1.8级;感白叶枯病(Ⅳ型菌5~7级,Ⅴ型5~7级)。
3.4 广泰优1002该组合是利用香型优质不育系广泰A与广恢1002组配而成的弱感光型迟熟三系杂交水稻品种。2017年参加华南感光晚籼组联合体区域试验,平均产量7.37 t/hm2,比对照博优998增产6.78%;2018年续试,平均产量7.53 t/hm2,比对照博优998增产4.76%;两年区域试验平均产量7.45 t/hm2,比对照博优998增产5.75%;2018年生产试验的平均产量为7.84 t/hm2,比对照博优998增产5.98%。在华南作双季晚稻种植,全生育期113.8 d,比对照博优998晚熟1 d。株高107.3 cm,穗长21.6 cm,667 m2有效穗数16.75万条,每穗总粒数155.8粒,结实率80.7%,千粒质量26.35 g。稻瘟病综合指数两年分别为3.1和5.5,穗颈瘟损失率最高达7级,白叶枯病7级,褐飞虱9级,稻米品质达部优2级,2020年通过国家品种审定(国审稻20200260)。
3.5 荣3优1002该组合是利用聚合了稻瘟病抗性基因Pi1、Pi2和白叶枯病抗性基因Xa23的双抗不育系荣3A与广恢1002组配选育而成的弱感光型迟熟三系杂交稻组合。稻瘟病全群抗性频率为92.9%~93.10%,对中B群、中C群的抗性频率分别为90.48%~100% 和100%,病圃鉴定叶瘟1.2~1.7级、穗瘟1.0~1.4级;对白叶枯病Ⅳ型和Ⅴ型菌的抗性级别均为1级,综合评价为高抗稻瘟和抗白叶枯。2017、2018年晚造参加广东省区试,平均产量分别为7.07、7.22 t/hm2,比对照广8优169分别增产10.84%、10.61%,增产均达极显著水平,增产点比例均为100%。2018年晚造参加广东省生产试验,平均产量为7.21 t/hm2,比对照广8优169增产9.01%。2019年通过广东省品种审定(粤审稻20190078)。
3.6 新泰优1002该组合是利用携带有稻瘟病抗性基因Pi1的优质不育系新泰A与广恢1002组配而成,在广东晚造全生育期114~115 d,比对照广8优169短1 d,比对照吉丰优1002短4 d。株型中集,分蘖力中等,耐寒性中等,抗倒力强。株高94.5~100.9 cm,667 m2有效穗数16.4万~18.1万,穗长20.3~21.1 cm,每穗总粒数143~144粒,结实率85.8%~86.4%,千粒质量24.3~24.9 g。稻瘟病全群抗性频率为78.6%~84.8%,对中B群、中C群的抗性频率分别为84.2%~90% 和80%~100%,病圃鉴定叶瘟1.4~2.0级、穗瘟1.0~3.0级;对白叶枯病Ⅳ、Ⅴ型菌抗性分别为3~5级和5~7级;米质鉴定为部标优质2级。2018、2019年晚造参加广东省区试,平均产量分别为6.56、7.83 t/hm2,比对照广8优169分别减产0.6% 和1.14%,减产均未达显著水平。2019年晚造参加广东省生产试验,平均产量为8.14 t/hm2,比对照吉丰优1002增产1.45%。
3.7 恒丰优1002该组合是利用优质不育系恒丰A与广恢1002组配育成,在华南作双季晚稻种植,全生育期117.6 d,比对照博优998晚熟4.3 d。株高106.8 cm,穗长22.6 cm,667 m2有效穗数16.5万,每穗总粒数170.6粒,结实率80.7%,千粒质量25.9 g。稻瘟病综合指数两年分别为3.9、5.2,穗颈瘟损失率最高达7级,白叶枯病5级,褐飞虱9级。整精米率60.4%,垩白度3.2%,直链淀粉含量16.2%,胶稠度70.0 mm,碱消值5.8级,长宽比3.3,达到农业行业标准《食用稻品种品质》 3级。2017年参加华南感光晚籼组联合体区域试验,平均产量为7.14 t/hm2,比对照博优998增产3.41%;2018年续试,平均产量为7.80 t/hm2,比对照博优998增产9.53%;两年区域试验平均产量为7.47 t/hm2,比对照博优998增产6.52%;2019年生产试验平均产量为8.06 t/hm2,比对照博优998增产12.21%。
4 广恢1002系列杂交稻的高产制种技术 4.1 精心选择适宜的制种区域,确保安全生产选择制种田时,须选择在土壤肥沃、前作最好未种植过水稻、田里无落粒谷、隔离条件好的地方进行制种。如果前作是水稻,则在制种翻耕前(30 d以上)对田地进行泡水,使落田谷发芽长秧,制种插秧前将秧苗翻耕打烂,以确保不影响制种。此外,如果选择的制种区周边有水稻种植,则制种田与大田生产水稻田块间隔距离不少于250 m,或者开花间隔时间不少于20 d,以避免其他水稻的花粉污染。
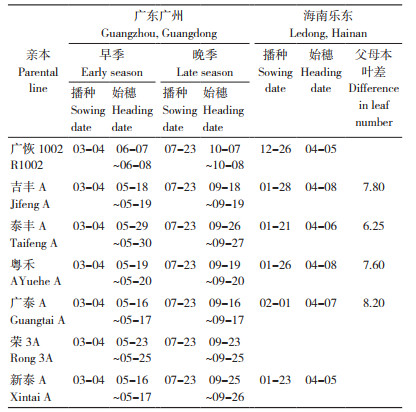
4.2 准确安排父母本播差期,确保花期相遇父母本花期相遇良好是杂交稻制种获得高产的重要基础。根据亲本的生育期及制种区域,合理安排亲本的播种期。吉丰优1002、粤禾优1002、泰优1002和广泰优1002的亲本在广东省广州市和海南省乐东县的生育期如表 3所示。在海南春制,第一期父本广恢1002一般安排在12月下旬播种,第二期父本比第一期父本迟播7~8 d;母本吉丰A、粤禾A、泰丰A和广泰A在次年1月下旬播种。父母本错期主要以叶差为主、时差为辅,一般父本比母本早播3~4 d为宜。
|
4.3 培育壮秧,为制种夺得高产奠定基础
种子要稀播匀播。父本的播种量为7.5~ 8.0kg/hm2,母本播种量为37.5~45.0 kg/hm2,秧龄在15~20 d为宜。秧苗生长期间需要加强肥水管理,秧田翻耕前施复合肥300 kg/hm2作基肥,3叶期和5叶期分别追施30、45 kg/hm2尿素,插秧前4~5 d喷施“送嫁药”,防止秧苗病虫害带入本田。
4.4 合理确定父母本行比安排合理的父母本行比是制种获得高产的关键之一。一般父母本行比以2 ∶ 10~11为宜,厢宽2.0~2.3 m。父本分蘖强,花期长,可偏施钾肥,培育穗大、花期长的健壮群体;吉丰A、粤禾A、泰丰A和广泰A这4个母本的生育期均相对较短,分蘖力一般,可适当增加种子用量,一般用不育系种子33.75~37.5 kg/hm2;父本用7.5 kg/hm2,确保插足基本苗,培育高产的健康群体。一般父本和母本每穴均插2~3苗。
4.5 科学管理水肥,构建高产制种群体施肥应遵循前重后轻的原则,以底肥为主、追肥为辅,后期视群体生长情况补肥,同时根据父母本的特性酌情调整。移栽前施复合肥和尿素各150 kg/hm2作基肥;移栽后5~7 d追施第1次肥,施尿素150 kg/hm2和复合肥150~225 kg/hm2;插秧后15 d追施第2次肥,分蘖期施复合肥600 kg/hm2和尿素150 kg/hm2。之后晒田3~5 d,晒田时宜施用除草剂如氰氟草酯,2 d后灌水,可有效杀灭稗草。幼穗分化期根据苖情追施第3次壮胎肥,即在每穴有2~3个苞时追施复合肥约225 kg/hm2。此外,根据田块差异、父母本错期、群体生长情况进行相应调整。
4.6 父母本发育进度检查与花期调节父母本进入幼穗分化期时要定期检查幼穗发育进度,根据父母本主穗的分化程度预测花期是否相遇。若花期相遇好,则继续进行常规观察及田间种植管理;若花期相遇不合理,则需通过肥水管控或喷施多效唑等进行花期调节。
4.7 适时适量喷施“九二〇”广恢1002系列组合的父母本对“九二〇”均较为敏感,在母本始穗10%~15% 时喷施以调节长势。第1、2次喷施“九二〇”的用量为180、90 g/hm2,且父母本同时喷施,两次喷施时间间隔1 d。株高以父本高于母本15~20 cm为宜,若父本低于母本或高于母本不足15 cm,则可以对父本再次少量喷施“九二〇”。
4.8 田间除杂保纯制种期间,从秧苗期、抽穗期、齐穗期到收割前,均须做好田间纯度调查和除杂工作,严格防止种子混杂,特别是抽穗期喷施“九二〇”前,要对父母本进行严格除杂保纯工作,确保种子质量。
4.9 适时收获,保质保量为确保制种群体有良好的通风透光环境,在母本授粉结束后立即割除父本,保障种子质量和纯度。杂交种子要做到适时收割,收割最好选择在晴天,防止收割时种子湿度过高,若不及时摊开晾晒将会导致种子出芽率下降。此外,收割前还需要注意彻底清洗收割机。翻晒种子时要彻底清扫地面和各种用具,避免机械混杂,最好直接用烘干机烘干种子。
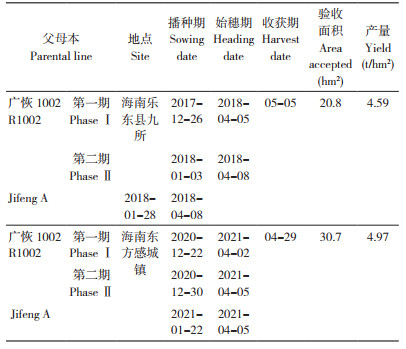
按照以上制种技术规程,创造了吉丰优1002等组合在海南的高产制种记录(表 4)。2018年广东华茂高科种业有限公司在海南乐东九所制种20.8 hm2,经实割验收,平均产量达到4.59 t/hm2;2021年该公司在海南东方感城镇制种30.7 hm2,创造了4.97 t/hm2的高产制种记录。
5 讨论与展望 5.1 杂交稻白叶枯病抗性育种的策略
水稻白叶枯病是由Xanthomonas oryzae pv.oryzae(Xoo)引起的细菌性病害,可导致水稻产量损失15%~25%,严重时可达50%[10],种植抗病品种是防治水稻白叶枯病最有效的方法。因此,在台风雨频繁、白叶枯病多发的华南沿海地区,抗白叶枯病是华南稻区水稻品种(尤其是晚造迟熟品种)所必须具备的一个重要性状。自20世纪60年代末以来,科学家们就致力于水稻白叶枯病抗性品种基因的发掘和利用。
近30年来、分子生物学技术飞速发展,为水稻抗白叶枯病基因资源的挖掘、基因鉴定、定位和克隆发挥了重要的促进作用。迄今已有46个显性和隐性的白叶枯病抗性基因被鉴定[11]、16个基因被克隆,包括Xa1[12]及其等位基因Xa2、Xa14、Xa31(t)、Xa45(t)[13-14]、Xa3/Xa26[15-16]、Xa4[17]、xa5[18-19]、Xa7[20]、Xa10[21]、xa13[22]、Xa21[23]、Xa23[24]、xa25[25]、Xa27[26]和xa41(t)[27]。在白叶枯病抗病基因中,杂交稻育种上以采用具有持久广谱、抗性的显性基因最好。根据上述已克隆基因的抗性机理和特点,发现Xa7和Xa23均属于执行者抗性基因(Eexecutor R gene),表现为显性遗传,且抗谱广[20, 24-28],是现有抗性基因中最适合应用于杂交稻育种的两个基因。其中,Xa7在连续多年的大田生产实践中证明,其抗性的持久性和耐高温能力也很强,且其持久抗白叶枯病能力优于Xa4和Xa10[28];此外,Xa7对广东近年来新出现的白叶枯Ⅸ型菌具有良好的抗性(数据未发表)。因此,在温度较高且新菌群不断演化的华南稻区,Xa7无疑是一个用于培育抗白叶枯病杂交稻较为理想的抗性基因。本研究利用Xa7育成的抗白叶枯病恢复系广恢312、广恢1002等,已组配育成天优312、博Ⅱ优312、吉丰优1002、泰优1002等系列杂交稻,从2007年博Ⅱ优312审定并投入生产应用开始到现在的吉丰优1002、泰优1002、广泰优1002等,Xa7已连续在华南稻区大面积应用15年,一直表现出很好的抗性,这进一步证明了其抗性持久的特点。但值得注意的是,利用携带Xa7的恢复系与感白叶枯病的不育系杂交,组配育成Xa7基因杂合的杂交稻(如吉丰优1002),在人工接种鉴定的条件下往往其白叶枯病抗性级别为中感或感(5~7级),达不到抗级,但在生产上的抗性多年来却一直表现很好,说明人工接种抗性与大田抗性存在一定差异。当利用携带Xa7的恢复系与携带Xa23基因的不育系杂交组配育成的杂交稻(如荣3优1002),其人工接种抗性和大田抗性均达到抗级水平(1级)。Xa23是从华南普通野生稻中挖掘到的显性广谱白叶枯病抗性基因,它的一个重要特点是全生育期抗病,可在苗期鉴定[29]。但据朱小源等接种鉴定发现,Xa23不抗华南地区新出现的白叶枯病优势菌群Ⅸ型菌(数据未发表)。
由此可见,在杂交稻白叶枯病抗性育种中,将Xa7和Xa23分别导入杂交稻的父本和母本中,培育同时聚合两个显性抗白叶枯病基因的杂交稻,不仅可以实现人工接种鉴定抗性与大田抗性表现高度一致,而且可以提高杂交稻白叶枯病抗性的抗谱,以及解决全生育期的抗病性问题。
5.2 弱感光型迟熟恢复系选育方法与育种效率水稻互作感光特性在遗传上受两对分别位于第7号和第8号染色体上的非等位基因LH1 (LP1) 和LH2 (LP2) 互作产生[6, 8-9]。一般的恢复系材料中,普遍携带LH1基因。因此,在育种实践上,培育华南互作感光型迟熟杂交稻可分为两条技术路线:一是选育携带有LH2基因的早中熟不育系(如博A、秋A),然后与携带有LH1的恢复系杂交,组配育成感光迟熟杂交稻;二是将LH1与LH2整合在一起,培育互作感光型迟熟恢复系(如广恢R3550、广恢55),与不育系组配,进而育成互作感光型杂交稻组合[6]。由于涉及到两个基因的互作,且只有两个基因同时共存的情况下,互作感光迟熟的表型才表现出来,因此,利用传常规的系统选育方法培育仅仅携带LH2的中早熟不育系(如博A、秋A等),必须要在选育保持系过程中,同时通过与携带LH1的不育系测配以观察其所选优良单株是否携带LH2基因,否则该基因在纯合稳定的各世代选种过程中很容易丢失。相对不育系的选育,利用常规育种方法培育同时携带LH1和LH2的恢复系则简单得多,往往只需选择生育期足够长的迟熟优良单株即可。为提高互作感光杂交稻亲本的育种选择效率,最好的方法就是利用分子标记辅助选择,以提高目标优良单株的选择准确性和选择效率,尤其是在培育既具有互作感光特性,又携带有其他优良性状(如抗稻瘟、抗白叶枯、抗虫或其他优良品质等)基因时,借助分子标记辅助选择显得尤为重要。本研究中,携带有Xa7的互作感光型迟熟恢复系广恢1002,就是综合应用了常规育种技术与分子标记辅助选择技术育成的。因此,加强常规育种与分子育种技术的结合,是提高杂交稻高产稳产、优质高效、绿色安全杂交稻育种效率的重要技术途径。
| [1] |
袁隆平. 中国的杂交水稻[J]. 中国水稻科学, 1986(1): 8-18. DOI:10.16819/j.1001-7216.1986.01.002 YUAN L P. Hybrid rice in China[J]. Chinese Journal of Rice Science, 1986(1): 8-18. DOI:10.16819/j.1001-7216.1986.01.002 |
| [2] |
任光俊, 颜龙安, 谢华安. 三系杂交水稻育种研究的回顾与展望[J]. 科学通报, 2016, 61(35): 3748-3760. DOI:10.1360/N972016-01109 REN G J, YAN L A, XIE H A. Retrospective and perspective on indica three-line hybrid rice breeding research in China[J]. Chinese Science Bulletin, 2016, 61(35): 3748-3760. DOI:10.1360/N972016-01109 |
| [3] |
彭惠普, 李维明, 伍应运, 黄德娟, 符福鸿, 王丰. 广谱恢复系3550及其系列杂交稻的选育和应用Ⅰ. 恢复系3550的选育及其特征特性[J]. 杂交水稻, 1993(6): 1-3. DOI:10.16267/j.cnki.1005-3956.1993.06.001 PENG H P, LI W M, WU Y Y, HUANG D J, FU F H, WANG F. Selection and utilization of rice restorer 3550 and its hybrid combinations I. Selection of restorer 3550 and its characteristics[J]. Hybrid Rice, 1993(6): 1-3. DOI:10.16267/j.cnki.1005-3956.1993.06.001 |
| [4] |
彭惠普, 李维明, 伍应运, 黄德娟, 符福鸿, 王丰. 广谱恢复系3550及其系列杂交稻的选育和应用Ⅱ. 3550系列杂交稻组合的试种及推广应用[J]. 杂交水稻, 1994(1): 9-11. DOI:10.16267/j.cnki.1005-3956.1994.01.003 PENG H P, LI W M, WU Y Y, HUANG D J, FU F H, WANG F. Selection and utilization of rice restorer 3550 and its hybrid combinations Ⅱ. Experimental demonstration and extension of hybrid combinations of 3550[J]. Hybrid Rice, 1994(1): 9-11. DOI:10.16267/j.cnki.1005-3956.1994.01.003 |
| [5] |
王丰. 杂交水稻育种成就与展望——广东省农业科学院杂交水稻研究50年回顾[J]. 广东农业科学, 2020, 47(12): 1-11. DOI:10.16768/j.issn.1004-874X.2020.12.001 WANG F. Achievements and prospects of hybrid rice breeding-Review of 50 years'research on hybrid rice by Rice Research Institute of Guangdong Academy of Agricultural Sciences[J]. Guangdong Agricultural Sciences, 2020, 47(12): 1-11. DOI:10.16768/j.issn.1004-874X.2020.12.001 |
| [6] |
李彦婷, 李金华, 柳武革, 付崇允, 廖亦龙, 朱满山, 刘振荣, 刘迪林, 曾学勤, 王丰. 弱感光型杂交稻吉丰优3550抽穗期性状QTL分析[J]. 分子植物育种, 2016(10): 2686-2695. DOI:10.13271/j.mpb.014.002686 LI Y T, LI J H, LIU W G, FU C Y, LIAO Y L, ZHU M S, LIU Z R, LIU D L, ZENG X Q, WANG F. Analysis on the quantitative trait locus for the heading date in the weakly photoperiod sensitive and late-maturing hybrid rice Jifeng You R3550[J]. Molecular Plant Breeding, 2016(10): 2686-2695. DOI:10.13271/j.mpb.014.002686 |
| [7] |
周长海, 王宝和, 赵步洪, 戴正元, 张洪熙. 从丰优香占的育成谈优质杂交籼稻育种[J]. 杂交水稻, 2007, 22(2): 11-13. DOI:10.16267/j.cnki.1005-3956.2007.02.003 ZHOU C H, WANG B H, ZHAO B H, DAI Z Y, ZHANG H X. Discussions on superior grain quality breeding of indica hybrid rice from the development of Fengyou Xiangzhan[J]. Hybrid Rice, 2007, 22(2): 11-13. DOI:10.16267/j.cnki.1005-3956.2007.02.003 |
| [8] |
李金华. 水稻互作感光和香味性状的遗传分析和基因定位研究[D]. 南昌: 江西农业大学, 2005. LI J H. Genetics analysis and gene mapping about interaction photoperiod sensitivity and fragrance traits of rice (Oryza sativa L. )[D]. Nanchang: Jiangxi Agricultural University, 2005. |
| [9] |
LIU S, WANG F, GAO L J, LI J H, LI R B, GAO H L, DENG G F, YANG J S, LUO X J. Genetic analysis and fine mapping of LH1 and LH2, a set of complementary genes controlling late heading in rice (Oryza sativa L.)[J]. Breeding Science, 2012, 62: 310-319. DOI:10.1270/jsbbs.62.310 |
| [10] |
ARSHAD H M I, SALEEM K, KHAN J A, RASHID M, ATIQ M, ALAAM S S, SAHI S T. Pathogenic diversity of Xanthomonas oryzae pv. oryzae isolates collected from Punjab Province of Pakistan[J]. European Journal of Plant Pathology, 2017, 147: 639-651. DOI:10.1007/s10658-016-1032-5 |
| [11] |
CHEN S, WANG C, YANG J, CH EN, B, WA NG, W, SU J, FENG A, ZENG L, ZHU X. Identification of the novel bacterial blight resistance gene Xa46(t) by mapping and expression analysis of the rice mutant H120[J]. Scientific Reports, 2020, 10: 12642. DOI:10.1038/s41598-020-69639-y |
| [12] |
YOSIMURA S, YAMANOUCHI U, KATAYOSE Y, TOKI S, WANG Z, KONO I, KURATA N, YANO M, SASAKI T. Expression of Xa1, a bacterial blight-resistance gene in rice, is induced by bacterial inoculation[J]. Proceedings of the National Academy of Sciences of the United States of America, 1998, 95: 1663-1668. DOI:10.1073/pnas.95.4.1663 |
| [13] |
JI C, JI Z, LIU B, CHENG H, LIU H, LIU S, YANG B, CHEN G. Xa1 allelic R genes activate rice blight resistance suppressed by interfering TAL effectors[J]. Plant Communications, 2020(1): 100087. DOI:10.1016/j.xplc.2020.100087 |
| [14] |
ZHANG B, ZHANG H, LI F, OUYANG Y, YUAN M, LI X, XIAO J, WANG S. Multiple alleles encoding atypical NLRs with unique central tandem repeats in rice confer resistance to Xanthomonas oryzae pv. oryzae[J]. Plant Communications, 2020(1): 100088. DOI:10.1016/j.xplc.2020.100088 |
| [15] |
SUN X, CAO Y, YANG Z, XU C, LI X, WANG S, ZHANG Q. Xa26, a gene conferring resistance to Xanthomonas oryzae pv. oryzae in rice, encodes an LRR receptor kinase-like protein[J]. Plant Journal, 2004, 37: 517-527. DOI:10.1046/j.1365-313x.2003.01976.x |
| [16] |
XIANG Y, CAO Y, XU C, LI X, WANG S. Xa3, conferring resistance for rice bacterial blight and encoding a receptor kinase-like protein, is the same as Xa26[J]. Theoretical Applied Genetics, 2006, 113: 1347-1355. DOI:10.1007/s00122-006-0388-x |
| [17] |
HU K, CAO J, ZHANG J, XIA F, KE Y, ZHANG H, XIE W, LIU H, CUI Y, CAO Y, SUN X, XIAO J, LI X, ZHANG Q, WANG S. Improvement of multiple agronomic traits by a disease resistance gene via cell wall reinforcement[J]. Nature Plants, 2017(3): 17009. DOI:10.1038/nplants.2017.9 |
| [18] |
IYER A S, MCCOUCH S R. The rice bacterial blight resistance gene xa5 encodes a novel form of disease resistance[J]. Molecular Plant-Microbe Interaction, 2004, 17: 1348-1354. DOI:10.1094/MPMI.2004.17.12.1348 |
| [19] |
JIANG G, XIA Z, ZHOU Y, WAN J, LI D, CHEN R, ZHAI W, ZHU L. Testifying the rice bacterial blight resistance gene xa5 by genetic complementation and further analyzing xa5 (Xa5) in comparison with its homolog TFⅡAγ1[J]. Molecular Genetics and Genomics, 2006, 275: 354-366. DOI:10.1007/s00438-005-0091-7 |
| [20] |
CHEN X, LIU P, MEI L, HE X, CHEN L, LIU H, SHEN S, JI Z, ZHENG X, ZHANG Y, GAO Z, ZENGG D, QIAN Q, MA B. Xa7, a new executor R gene that confers durable and broad-spectrum resistance to bacterial blight disease in rice[J]. Plant Communications, 2021(2): 100143. DOI:10.1016/j.xplc.2021.100143 |
| [21] |
TIAN D, WANG J, ZENG X, GU K, QIU C, YANG X, ZHOU Z, GOH M, LUO Y, MURATA-HORI M, WHITE F F, YIN Z. The rice TAL effector-dependent resistance protein XA10 triggers cell death and calcium depletion in the endoplasmic reticulum[J]. Plant Cell, 2014, 26: 497-515. DOI:10.1105/tpc.113.119255 |
| [22] |
YANG B, SUGIO A, WHITE F. Os8N3 is a host disease-susceptibility gene for bacterial blight of rice[J]. Proceedings of the National Academy of Sciences of the United States of America, 2006, 103: 10503-10508. DOI:10.1073/pnas.0604088103 |
| [23] |
SONG W, WANG G, CHEN L, KIM HS, PI L, HOLSTEN T, GARDNER J, WANG B, ZHAI W, ZHU L, FAUQUET C, RONALD P. A receptor kinase-like protein encoded by the rice disease resistance gene, Xa21[J]. Science, 1995, 270: 1804-1806. DOI:10.1126/science.270.5243.1804 |
| [24] |
WANG C, ZHANG X, FAN Y, GAO Y, ZHU Q, ZHENG C, QIN T, LI Y, CHE J, ZHANG M, YANG B, LIU Y, ZHAO K. XA23 is an executor R protein and confers broad-spectrum disease resistance in rice[J]. Molecular Plant, 2015, 8: 290-302. DOI:10.1016/j.molp.2014.10.010 |
| [25] |
LIU Q, YUAN M, ZHOU Y, LI X, XIAO J, WANG S. A paralog of the MtN3/saliva family recessively confers race-specific resistance to Xanthomonas oryzae in rice[J]. Plant Cell Environment, 2011, 34: 1958-1969. DOI:10.1111/j.1365-3040.2011.02391.x |
| [26] |
GU K, YANG B, TIAN D, WU L, WANG D, SREEKALA C, YANG F, CHU Z, WANG G L, WHITE F. R gene expression induced by a type-Ⅲ effector triggers disease resistance in rice[J]. Nature, 2005, 435: 1122-1125. DOI:10.1038/nature03630 |
| [27] |
HUTIN M, SABOT FO, GHESQUIERE A, KOEBINK R, SZUREK B. A knowledge-based molecular screen uncovers a broad-spectrum OsSWEET14 resistance allele to bacterial blight from wild rice[J]. Plant Journal, 2016, 84: 694-703. DOI:10.1111/tpj.13042 |
| [28] |
VERA CRUZ C M, BAI J, ONA I, LEUNG H, NELSON R J, MEW T W, LEACH J E. Predicting durability of a disease resistance gene based on an assessment of the fitness loss and epidemiological consequences of avirulence gene mutation[J]. Proceedings of the National Academy of Sciences of the United States of America, 2000, 97: 13500-13505. DOI:10.1073/pnas.250271997 |
| [29] |
章琦. 水稻白叶枯病抗性基因鉴定进展及其利用[J]. 中国水稻科学, 2005, 19(5): 453-459. DOI:10.16819/j.1001-7216,2005.05.012 ZHANG Q. Highlights in identification and application of resistance genes to bacterial blight[J]. Chinese Journal of Rice Science, 2005, 19(5): 453-459. DOI:10.16819/j.1001-7216,2005.05.012 |
(责任编辑 崔建勋)