 2023, Vol. 50
2023, Vol. 50文章信息
引用本文 |

基金项目
- 国家自然科学基金(32101647);仲恺农业工程学院仲华基因产业学院项目(ZHCY2023007)
作者简介
- 蔡欣(1998—),女,在读硕士生,研究方向为水稻重要农艺性状的分子机理,E-mail:254767327@qq.com.
通讯作者
- 王丽敏(1984—),女,博士,副教授,研究方向为水稻重要农艺性状的分子机理,E-mail:wanglimin@zhku.edu.cn.
文章历史
- 收稿日期:2023-10-25

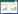


【研究意义】转录因子(Transcription factor, TF)可在细胞核中直接与靶标基因启动子的特异性顺式元件结合,形成具有RNA聚合酶活性的转录复合体,从而抑制或激活下游基因的转录;还可与顺式元件相互作用来调节特定基因的表达、响应环境胁迫[1-2]。bHLH(Basic Helix-Loop-Helix)转录因子家族是植物中仅次于MYB家族的第二大家族[3],在植物的生长发育过程中具有重要作用,其家族成员的功能尚未完全解晰。【前人研究进展】bHLH转录因子具有高度保守的bHLH结构域,该保守结构域由约60个保守氨基酸残基和2个保守基序组成,一个保守基序为基础结构域(Basic region),位于N端,约15个氨基酸;另一个为HLH(Helix-Loop Helix)结构域,位于C端,由约50~60个保守氨基酸组成[4-5]。不同植物中的bHLH基因家族成员数量不同,在拟南芥中发现162个转录因子家族成员[6]、水稻中发现167个[7]、玉米中发现203个[8]、红花中发现41个[1]。bHLH蛋白可调节许多生理过程,在信号转导[9]、植物生长发育[10-12]和铁稳态调控[13]等方面发挥重要作用。拟南芥ICE1编码一种MYC-like的bHLH转录因子,可以与CBF3启动子序列特异性结合,在过表达ICE1的植株中,低温条件下CBF基因表达增强,提高了拟南芥的抗寒能力[14];黄瓜ICE1可响应并激活CBF3和COR的表达,从而提高黄瓜的抗寒能力[15-16];在小麦中,TabHLH13的表达量在盐胁迫条件下迅速上升,以调控小麦的抗盐碱能力[17];在番茄中,MYB和bHLH蛋白共同作用会影响花青素的合成[18]。水稻bHLH转录因子在水稻抗旱、抗寒等过程中发挥重要作用,OsbHLH148可通过参与茉莉酸(JA)信号途径调节水稻的抗旱性[19];OsbHLH120可通过参与ABA信号通路,使植株在干旱胁迫下仍能保持一定的含水量,从而提高水稻的抗旱能力[20-21]。Zhang等[22]研究发现,bHLH57通过正调控海藻糖合成、活性氧代谢和CBF/DREB依赖途径来增强水稻耐冷性。bHLH转录因子还可与MYB转录因子互作,形成复杂的调控网络调控植物生长发育进程。Lee等[23]发现OsbHLH073通过调节与OsKO1、OsKO2基因相关的GA生物合成,参与调控株高、节间伸长和穗外露等性状。铁是植物生长发育所必需的元素,已有许多研究表明bHLH转录因子参与铁稳态的维持;OsPRI1(OsbHLH060)为正调控铁稳态的转录因子,能调控许多缺铁响应基因的表达[24];OsbHLH061作用于OsPRI1上游,是多个铁稳态相关基因表达的负调控因子[25]。
【本研究切入点】bHLH转录因子在植物生长发育、逆境响应及激素信号传导方面发挥重要作用,水稻中存在众多bHLH转录因子家族成员,但其功能尚不明确。开展水稻bHLH转录因子家族基因的鉴定及表达分析,能为进一步研究其功能、解析其调控机理提供理论基础。【拟解决的关键问题】本研究鉴定得到1个编码bHLH转录因子的基因OsbHLH109(LOC_Os01g67480),通过生物信息学技术分析该基因的理化性质、基因结构、构建系统进化树;利用qRT-PCR技术检测不同条件下OsbHLH109的表达模式,旨在探讨以下问题:(1)OsbHLH109的蛋白结构及进化关系;(2)正常条件下OsbHLH109的表达模式;(3)OsbHLH109在激素及非生物胁迫条件下的表达模式。为深入研究OsbHLH109转录因子在植物抗逆性调控中的作用机制提供理论支持和研究方向。
1 材料与方法 1.1 试验材料供试水稻材料为粳稻品种中花11(ZH11)。
1.2 生物信息学分析利用NCBI数据库和国家水稻数据库(https://www.ricedata.cn/gene/)获取OsbHLH109的编码区及氨基酸序列,通过Smart在线软件(http://smart.embl-heidelberg.de/)对OsbHLH109蛋白结构域进行预测分析,通过Blast分析在Uniprot数据库(https://www.uniprot.org/)中找到其他物种中与OsbHLH109同源性最高的同源蛋白序列,利用MEGA 11.0的ClustalW软件对OsbHLH109及其同源蛋白进行氨基酸序列多重比对,并使用近邻法构建系统进化树。将起始密码子ATG上游2 000 bp的基因组序列设置为该基因的启动子序列。利用New PLACE在线分析软件(https://www.dna.affrc.go.jp/PLACE/?action=newplace)对启动子序列进行顺式作用元件分析。
1.3 试验方法1.3.1 表达模式分析 于2023年3月萌发水稻种子,待种子萌发后,将水稻幼苗置于光照培养箱中(28 ℃光照培养12 h、25 ℃黑暗培养12 h,相对湿度60%),使用木村B营养液(北京酷莱搏科技有限公司,NSP1020)培养[26],每隔3 d更换1次营养液。培养至3~4叶期时,提取ZH11幼根的总RNA;同时将萌发后的ZH11播种于广东省广州市试验田中。在孕穗期时,提取ZH11茎、叶和幼穗(2~3 cm)的总RNA用于组织特异性表达分析。以上每个处理均3次重复。
1.3.2 植物激素处理及取样 参照1.3.1方法将水稻幼苗培养至3~4叶期后,分别移至含有10-5 mol/L各植物激素的木村B营养液中进行处理。试验用激素为:细胞分裂素(6-Benzylaminopurine,6-BA,上海麦克林生化科技股份有限公司生产,分析纯试剂)、生长素(Indole acetic acid,IAA,北京酷来搏科技有限公司生产,分析纯试剂)、赤霉素(Gibberellic acid,GA3,罗恩麦克林阿拉丁Sigma国药厂生产,分析纯药剂)、脱落酸(Abscisic acid,ABA,北京酷来搏科技有限公司生产,分析纯试剂)[27-28]。其他培养条件与1.3.1一致,分别于激素处理0、1、2、8、24 h时取水稻叶片组织样品并提取总RNA,用于OsbHLH109的表达分析,每个处理3次重复。
1.3.3 非生物胁迫处理及取样 参照1.3.1方法将水稻幼苗培养至3~4叶期后,分别转移到含有0.2 mol/L NaCl(盐胁迫)、20% PEG 6000的水培营养液(干旱)及42 ℃(高温)、4 ℃(低温)的光照培养箱中进行处理,其他培养条件与1.3.1一致。分别于处理0、1、2、8、24 h时取水稻叶片组织样品并提取总RNA,用于OsbHLH109的表达分析,每个处理3次重复。
1.4 RNA提取和qRT-PCR扩增使用TRIzol试剂(Glpbio公司)提取水稻样品总RNA,使用逆转录试剂盒(南京诺唯赞生物科技股份公司)将1 µg总RNA反转录合成第一链cDNA。根据OsbHLH109基因序列,使用NCBI数据库中Primer-Blast在线软件设计特异引物,并于广州艾基生物技术有限公司合成,引物序列见表 1。以水稻Actin1为内参基因,qRT-PCR反应体系为20 µL,反应程序为95 ℃预变性3 min,1个循环;95 ℃变性5 s,60 ℃延伸30 s,40个循环。每个样品3次重复,采用2-ΔΔCT法计算OsbHLH109基因的相对表达量,通过单因素方差进行差异显著性分析,P<0.05表示差异显著。

2 结果与分析 2.1 OsbHLH109基因的结构分析
通过NCBI数据库、国家水稻数据库和Smart在线软件分析,结果表明OsbHLH109基因CDS全长为1 164 bp,包含6个外显子,共编码388个氨基酸;包含bHLH结构域,属于典型的bHLH转录因子,该基因编码的氨基酸序列N端第321~371个氨基酸为1个保守的HLH结构域。

|
| 灰色部分为3’UTR区及5’UTR区,黑色框为外显子,黑色细线为内含子,黑色数字为外显子碱基数 Gray parts are 3'UTR region and 5'UTR region, the black box is exon, the black thin line is intron, and the number of exon bases are presented by black numbers 图 1 OsbHLH109基因结构及蛋白结构域 Fig. 1 Gene structure model and the protein domain of OsbHLH109 |
2.2 OsbHLH109同源蛋白序列对比及进化分析
将OsbHLH109与其他植物的bHLH109同源蛋白进行氨基酸序列比对,结果显示OsbHLH109与其他同源蛋白在321~371个氨基酸的区域高度相似(图 2)。进一步系统进化树分析表明(图 3),OsbHLH109与黍(Panicum hallii, XP_025814593.1)、谷子(Setaria italica, XP_004970931.1)、小麦(Triticum aestivum, XP_044342445.1)、大麦(Hordeum vulgare, KAI5007066.1)的bHLH109同源蛋白有相对较近的演化关系;而与拟南芥(Arabidopsis thaliana, NP_192657.1)、玉米(Zea mays, PWZ34064.1)、芒草(Miscanthus lutarioriparius, CAD6343449.1)、高粱(Sorghum bicolor, XP_002456792.2)、二穗短柄草(Brachypodium distachyon, PNT73394.1)的bHLH109同源蛋白的演化关系较远。

|
| OsbHLH109:水稻(Oryza sativa, XP_015613853.1);PhbHLH109:黍(Panicum hallii, XP_025814593.1);SibHLH109:谷子(Setaria italica, XP_004970931.1);TabHLH109:小麦(Triticum aestivum, XP_044342445.1);HvbHLH109:大麦(Hordeum vulgare, KAI5007066.1);AtbHLH109:拟南芥(Arabidopsis thaliana, NP_192657.1);ZmbHLH109:玉米(Zea mays, PWZ34064.1);MlbHLH109:芒草(Miscanthus lutarioriparius, CAD6343449.1);SbbHLH109:高粱(Sorghum bicolor, XP_002456792.2);BdbHLH109:二穗短柄草(Brachypodium distachyon, PNT73394.1) 图 2 OsbHLH109及其同源蛋白的氨基酸序列比对 Fig. 2 Amino acid sequences alignment of OsbHLH109 and its homologous proteins |

|
| 图 3 OsbHLH109及其同源蛋白的进化关系分析 Fig. 3 Phylogenetic analysis of OsbHLH109 and its homologous proteins |
2.3 OsbHLH109启动子区域的顺式作用元件分析
利用New PLACE在线分析软件对OsbHLH109的ATG上游2 000 bp的序列进行顺式作用元件分析。结果发现,OsbHLH109的启动子区域存在17个光响应顺式作用元件、4个低氧胁迫顺式作用元件、2个水杨酸(Salicylic acid, SA)响应元件、10个ABA响应元件、2个GA响应元件、2个IAA响应元件、8个JA响应元件、1个防御与胁迫响应元件、2个MYB转录因子结合的相关元件、1个调控类玉米醇溶蛋白代谢的相关元件。
2.4 OsbHLH109基因的表达谱分析对中花11苗期的根、孕穗期的叶、茎和幼穗(2~3 cm)进行取样,检测水稻不同组织的OsbHLH109的转录表达水平发现,OsbHLH109在水稻根、茎、叶和幼穗中均有表达,其中叶部的表达水平相对最高(图 4),表明OsbHLH109的表达具有明显的时空特异性,可能在水稻不同组织部位发挥生物学功能。

|
| Actin1为内参基因,数据表示为平均值± SD,n = 3;** 表示与根部差异显著 Actin1 is an internal reference, and data are presented as the mean ± SD, n = 3;** represents significant difference compared with that of root 图 4 OsbHLH109在水稻不同组织中的表达水平分析 Fig. 4 Expression level analysis of OsbHLH109 in different tissues of rice |
2.5 外源激素处理对OsbHLH109基因表达的影响分析
OsbHLH109启动子区域存在若干个与激素响应相关的元件,推测OsbHLH109可能参与水稻激素响应,不同激素处理后水稻幼苗叶中OsbHLH109的表达水平如图 5所示。6-BA处理后,叶片中OsbHLH109的表达水平先降低后升高,在处理1 h时表达水平显著下调,24 h时表达水平相对最高,约为初始表达水平的4.11倍;IAA处理2 h时的表达水平显著升高,约为初始表达水平的17.29倍。GA3处理后,OsbHLH109的表达整体水平显著下调,处理2 h时表达水平相对最低,约为初始表达水平的31%。ABA处理后,OsbHLH109的表达整体水平显著高于初始水平,处理1 h时表达水平相对最高。表明OsbHLH109基因受以上4种激素诱导表达。

|
| Actin1为内参基因,数据为平均值± SD,n = 3;* 表示差异显著;** 表示差异极显著 Actin1 is an internal reference, and data are presented as the mean ± SD, n = 3; * represents significant difference; ** represents extremely significant difference 图 5 不同激素处理后OsbHLH109在水稻叶片中的表达水平分析 Fig. 5 Expression level analysis of OsbHLH109 in leaf tissues of rice after different hormone treatments |
2.6 非生物胁迫处理对OsbHLH109基因表达的影响分析
OsbHLH109启动子区域存在若干个与逆境响应相关的元件,推测OsbHLH109可能参与水稻逆境胁迫响应。对水稻幼苗分别进行低温(4 ℃)、干旱(PEG)、高温(42 ℃)和盐胁迫(NaCl)处理,检测水稻幼苗叶中OsbHLH109的表达水平(图 6)。低温处理后,水稻叶片中OsbHLH109其表达水平呈现先升后降再升的趋势,处理24 h时其表达水平最高。干旱处理后,表达水平在处理1 h时显著提高,约为初始表达水平的1.65倍。高温处理1 h后,表达水平整体显著上调,处理2 h时的表达水平达到峰值,约为初始表达水平的6.73倍。盐胁迫处理后,OsbHLH109的表达量呈现为先上升后下降再上升的趋势,在处理8 h时表达水平最高,约为初始表达水平的3.05倍。表明OsbHLH109基因的表达受低温、干旱、高温和盐胁迫诱导。

|
| Actin1为内参基因,数据为平均值± SD,n = 3;* 表示差异显著;** 表示差异极显著 Actin1 is an internal reference, and data are presented as the mean ± SD, n = 3; * represents significant difference; ** represents extremely significant difference 图 6 不同胁迫处理后OsbHLH109在水稻叶片中的表达水平分析 Fig. 6 Expression level analysis of OsbHLH109 in leaf tissues of rice after different stress treatments |
3 讨论
bHLH转录因子在植物的生长发育中起重要的作用[29]。本研究发现1个水稻bHLH转录因子基因家族成员OsbHLH109,其编码的氨基酸序列包含HLH结构域,属于典型的bHLH转录因子。OsbHLH109与其他植物中的同源蛋白表现出高度保守。系统进化分析表明,OsbHLH109蛋白与单子叶植物中同源蛋白具有更近的亲缘进化关系,与双子叶植物中同源蛋白具有相对较远的进化关系,形成了有明显差异的亚类分枝,推测OsbHLH109蛋白在物种进化过程中受到了选择。Li等[7]对水稻bHLH转录因子进行了系统进化分析,将OsbHLH家族的167个成员分为22个亚家族,结果表明,OsbHLH109与OsbHLH110、OsbHLH111、OsbHLH112和OsbHLH143演化关系较近。OsbHLH111被证实是与糖饥饿相关的生长抑制子,糖不足时,OsbHLH111的表达及其与蔗糖非发酵相关蛋白激酶OsSnRK1a的结合均增加,随后OsSnRK1a介导的OsbHLH111磷酸化增强了其与海藻糖6- 磷酸酶7(OsTPP7)启动子E-box的结合,从而增加了对OsTPP7的转录抑制,导致海藻糖6- 磷酸含量增加而蔗糖含量降低,最终抑制水稻生长[30]。启动子在基因的表达调控过程中起到关键作用,对未知功能的基因进行启动子序列分析可为进一步研究该基因的表达模式及生物学功能提供参考[31]。OsbHLH109启动子区域存在数个与水稻激素响应相关的顺式作用元件、若干与防御和逆境相关的调控元件和MYB转录因子结合位点相关等元件,说明OsbHLH109的转录表达可能受到逆境胁迫的调节,并参与各种逆境胁迫响应。前人研究表明,多数bHLH转录因子家族基因在植物不同组织器官中表达水平不同,而且其表达水平受一些环境条件的诱导,水稻中DPF编码基因(OsbHLH025)主要在根和穗中表达,稻瘟病菌侵染、氯化铜以及紫外光会诱导叶片中DPF表达[32]。OsSGI1(OsbHLH111)在水稻根中的表达量较高[30],磷缺乏时,OsPTF1(OsHLH096)在根部诱导特异性表达[33]。OsbHLH109的组织表达谱分析结果表明,OsbHLH109在水稻叶片组织中的表达水平均显著高于其他组织部位,推测OsbHLH109在水稻叶细胞活动中发挥重要功能。
大量研究表明,植物bHLH转录因子参与调节各种信号转导、生长发育及响应生物及非生物胁迫等重要途径[34]。在干旱胁迫下,RERT1(OsHLH006)通过JA信号途径参与干旱胁迫和创伤反应[35-36]。有研究筛选出22个干旱胁迫差异表达的bHLH家族基因并对候选基因进行比较分析,使用PEG模拟干旱胁迫时,大部分候选基因的PEG胁迫表达谱与干旱胁迫表达谱一致,而外源施加ABA时,大部分bHLH基因表达受到抑制,推测这些候选bHLH基因在响应干旱胁迫过程中受ABA负调控[37]。本研究中OsbHLH109的表达也受干旱和ABA诱导,表明OsbHLH109也可能通过ABA信号参与干旱胁迫响应。
转录因子通过控制结构基因的表达强度和方式,可有效地启动或关闭次生代谢合成途径[38]。有研究指出OsbHLH6(RERJ1)通过在细胞核与细胞质间的移动,动态调节SA和JA激素信号[39]。OsbHLH153和OsbHLH174对参与株型调控的油菜素内酯(Brassinolide,BR)与IAA响应迅速[40]。OsbHLH109的启动子区域也存在多个SA和JA响应元件,表明OsbHLH109也可能参与SA和JA信号调节。同时,植物激素响应表达分析结果显示,不同激素处理(ABA、GA3、IAA、6-BA)下,叶片中OsbHLH109的表达量均与对照达到显著差异水平,表明OsbHLH109可能参与多种激素调控。
bHLH转录因子还可与其他转录因子形成调控复合体协同调控植物的多种生命活动,盐胁迫是影响植物生长发育的重要非生物胁迫之一,严重制约农业生产和经济发展,转录因子在植物耐盐调控中起重要作用[41]。OsJAZ9作为转录因子,通过与OsNINJA和OsbHLH蛋白形成转录调控复合物控制JA合成通路来调控植株的耐盐性[42];OsbHLH120能够被PEG、盐和ABA强烈诱导[16];OsbHLH1基因在幼苗期能特异地被冷胁迫诱导表达[43];OsbHLH109的表达在盐害、低温、高温和干旱胁迫中出现显著变化,推测OsbHLH109可能通过多种信号途径参与水稻的逆境胁迫响应。
4 结论本研究明确了水稻bHLH转录因子基因OsbHLH109的基因结构、蛋白理化关系、蛋白结构域,发现OsbHLH109编码的氨基酸序列包含HLH结构域,属于典型的bHLH转录因子。OsbHLH109与其同源蛋白表现出高度保守,系统进化分析表明,与双子叶植物相比,OsbHLH109蛋白与单子叶植物中的同源蛋白的亲缘进化关系更近。OsbHLH109启动子区域存在数个与水稻激素响应相关的顺式作用元件、若干与防御和逆境相关的调控元件以及MYB转录因子结合位点相关等元件,表明OsbHLH109的转录表达可能受逆境胁迫调节,并参与各种逆境胁迫响应。组织表达谱分析表明,OsbHLH109在水稻叶片组织中高表达;不同激素处理(ABA、GA3、IAA、6-BA)下,叶片中OsbHLH109的表达量均与对照达到显著差异水平;不同胁迫处理(低温、高温、盐害、干旱)下,OsbHLH109的表达量出现显著变化,推测OsbHLH109可能通过ABA、GA3、IAA、6-BA等激素信号转导途径参与水稻的非生物胁迫响应。
| [1] |
HONG Y Q, NAVEED A, TIAN Y Y, LIU J Y, WANG L Y, WANG G, LIU X M, DONG Y Y, WANG F W, LIU W C, LI X W, ZHAO X, YAO N, LI H Y. Genome-wide identification, expression analysis, and subcellular localization of Carthamus tinctorius bHLH transcription factors[J]. International Journal of Molecular Sciences, 2019, 20(12): 3044. DOI:10.3390/ijms20123044 |
| [2] |
WANG J Y, HU Z Z, ZHAO T M, YANG Y W, CHEN T Z, YANG M L, YU W G, ZHANG B L. Genome-wide analysis of bHLH transcription factor and involvement in the infection by yellow leaf curl virus in tomato (Solanum lycopersicum)[J]. BMC Genomics, 2015, 16(1): 1-14. DOI:10.1186/s12864-015-1249-2 |
| [3] |
FELLER A, MACHEMER K, BRAUN E L, GROTEWOLD E. Evolutionary and comparative analysis of MYB and bHLH plant transcription factors[J]. The Plant Journal, 2011, 66(1): 94-116. DOI:10.1111/j.1365-313X.2010.04459.x |
| [4] |
TOLEDO-ORTIZ G, HUQ E, QUAIL P H. The Arabidopsis basic/Helix-loop-Helix transcription factor family[J]. Plant Cell, 2003, 15(8): 1749-1770. DOI:10.1105/tpc.013839 |
| [5] |
SHIMOMURA I, HAMMER R E, RICHARDSON J A, IKEMOTO S, BASHMAKOV Y, GOLDSTEIN J L, BROWN M S. Insulin resistance and diabetes mellitus in transgenic mice expressing nuclear SREBP-1c in adipose tissue: Model for congenital generalized lipodystrophy[J]. Genes & Development, 1998, 12(20): 3182-3194. DOI:10.1101/gad.12.20.3182 |
| [6] |
BAILETY P C, MARTIN C, TOLOEDO-ORTIZ G, QUAIL P H, HUQ E, HEIM M A, JAKOBY M, WERBER M, WEISSHAAR B. Update on the basic Helix-Loop-Helix transcription factor gene family in Arabidopsis thaliana[J]. Plant Cell, 2003, 15(11): 2497-2501. DOI:10.1105/tpc.151140 |
| [7] |
LI X X, DUAN X P, JIANG H X, SUN Y J, TANG Y P, YUAN Z, GUO J K, LIANG W Q, CHEN L, YIN J Y, MA H, WANG J, ZHANG D B. Genome-wide analysis of basic/Helix-Loop-Helix transcription factor family in rice and Arabidopsis[J]. Plant Physiology, 2006, 141(4): 1167. DOI:10.1104/pp.106.080580 |
| [8] |
CHANG W H, LAI A G. Genome-wide analyses of the bHLH superfamily in crustaceans: Reappraisal of higher-order groupings and evidence for lineage specific duplications[J]. Royal Society Open Science, 2018, 5(3): 172433. DOI:10.1098/rsos.172433 |
| [9] |
ZHU E G, YOU C J, WANG S S, CUI J, NIU B X, WANG Y X, QI J, MA H, CHANG F. The DYT1-interacting proteins bHLH010, bHLH089 and bHLH091 are redundantly required for Arabidopsis anther development and transcriptome[J]. The Plant Journal, 2015, 83(6): 976-990. DOI:10.1111/tpj.12942 |
| [10] |
LU R, ZHANG J, LIU D, WEI Y L, WANG Y, LI X B. Characterization of bHLH /HLH genes that are involved in brassinosteroid (BR) signaling in fiber development of cotton (Gossypium hirsutum)[J]. BMC Plant Biology, 2018, 18(1): 304. DOI:10.1186/s12870-018-1523-y |
| [11] |
XING B C, YANG D F, YU H Z, ZHANG B X, YAN K J, ZHANG X M, HAN R L, LIANG Z S. Overexpression of SmbHLH10 enhances tanshinones biosynthesis in Salvia miltiorrhiza hairy roots[J]. Plant Science, 2018, 276: 229-238. DOI:10.1016/j.plantsci.2018.07.016 |
| [12] |
ZHU Z G, LIANG H L, CHEN G P, LI F F, WANG Y S, LIAO C G, HU Z L. The bHLH transcription factor SlPRE2 regulates tomato fruit development and modulates plant response to gibberellin[J]. Plant Cell Reports, 2019, 38(9): 1053-1064. DOI:10.1007/s00299-019-02425-x |
| [13] |
WANG S D, LI L, YING Y H, WANG J, SHAO J F, YAMAJI N, WHELAN J, MA J F, SHOU H X. A transcription factor OsbHLH156 regulates strategy Ⅱ iron acquisition through localising IRO2 to the nucleus in rice[J]. New Phytologist, 2020, 225(3): 1247-1260. DOI:10.1111/nph.16232 |
| [14] |
CHINNUSAMY V, OHTA M, KANRAR S, Lee B H, HONG X H, AGARWAL M, ZHU J K. ICE1:A regulator of cold-induced transcriptome and freezing tolerance in Arabidopsis[J]. Genes & Development, 2003, 17(8): 1043-1054. DOI:10.1101/gad.1077503 |
| [15] |
LIU L Y, DUAN L S, ZHANG J C, ZHANG Z, MI G, REN H. Cucumber (Cucumis sativus L.) over-expressing cold-induced transcriptome regulator ICE1 exhibits changed morphological characters and enhances chilling tolerance[J]. Scientia Horticultur ae, 2010, 124(1): 29-33. DOI:10.1016/j.scienta.2009.11.018 |
| [16] |
XIANG D J, HU X Y, ZHANG Y, YIN K D. Over-expression of ICE1 gene in transgenic rice improves cold tolerance[J]. Rice Science, 2008, 15(3): 173-178. DOI:10.1016/S1672-6308(08)60039-6 |
| [17] |
符思路, 刘国祥, 张立超, 夏川, 赵光耀, 贾继增, 孔秀英. 小麦盐胁迫相关基因TabHLH13的克隆与分析[J]. 植物遗传资源学报, 2014, 15(5): 1006-1011. DOI:10.13430/j.cnki.jpgr.2014.05.013 FU S L, LIU G X, ZHANG L C, XIA C, ZHAO G Y, JIA J Z, KONG X Y. Cloning and analysis of a salt stress related gene TabHLH13 in wheat[J]. Journal of Plant Genetic Resources, 2014, 15(5): 1006-1011. DOI:10.13430/j.cnki.jpgr.2014.05.013 |
| [18] |
OUTCHKOUROV N S, CAROLLO C A, GOMEZ-ROLDAN V, VOS R C, BOSCH D, HALL R D, BEEKWILDER J. Control of anthocyanin and non-flavonoid compounds by anthocyanin-regulating MYB and bHLH transcription factors in Nicotiana benthamiana leaves[J]. Frontiers in Plant Science, 2014, 5: 519. DOI:10.3389/fpls.2014.00519 |
| [19] |
SEO J S, JOO J, KIM M J, KIM Y K, NAHM B H, SONG S I, CHEONG J J, LEE J S, KIM J K, CHOI Y D. OsbHLH148 a basic helix-loop-helix protein interacts with OsJAZ proteins in a jasmonate signaling pathway leading to drought tolerance in rice[J]. Plant Journal for Cell & Molecular Biology, 2011, 65(6): 907-921. DOI:10.1111/j.1365-313X.2010.04477.x |
| [20] |
LI J Z, HAN Y C, LIU L, CHEN Y P, DU Y X, ZHANG J, SUN H Z, ZHAO Q Z. qRT9, a quantitative trait locus controlling root thickness and root length in upland rice[J]. Journal of Experimental Botany, 2015, 66(9): 2723-2732. DOI:10.1093/jxb/erv076 |
| [21] |
BABITHA K C, VEMANNA R S, NATARAJA K N, UDAYAKUMAR M. Overexpression of EcbHLH57 transcription factor from Eleusine coracana L. in tobacco confers tolerance to salt, oxidative and drought stress[J]. PLoS One, 2015, 10(9): e0137098. DOI:10.1371/journal.pone.0137098 |
| [22] |
ZHANG L, XIANG Z P, LI J F, WANG S Y, CHEN Y, LIU Y, MAO D D, LUAN S, CHEN L B. bHLH57 confers chilling tolerance and grain yield improvement in rice[J]. Plant Cell Environment, 2023, 46(4): 1402-1418. DOI:10.1111/pce.14513 |
| [23] |
LEE J, MOON S, JANG S, LEE S, AN G, JUNG K H, PARK S K. OsbHLH073 negatively regulates internode elongation and plant height by modulating GA homeostasis in rice[J]. Plants, 2020, 9(4): 547. DOI:10.3390/plants9040547 |
| [24] |
ZHANG H M, LI Y, YAO X N, LIANG G, YU D Q. Positive regulator of iron homeostasis1, OsPRI1, facilitates iron homeostasis[J]. Plant Physiology, 2017, 175(1): 543-554. DOI:10.1104/pp.17.00794 |
| [25] |
WANG W J, YE J, XU H, LIU X, FU Y, ZHANG H, ROUACHED H, WHELAN J, SHEN Z G, ZHENG L Q. OsbHLH061 links topless/topless-related repressor proteins with positive regulator of iron homeostasis 1 to maintain iron homeostasis in rice[J]. The New Phytologist, 2022(5): 234. DOI:10.1111/nph.18096 |
| [26] |
姜晓婷, 黄高翔, 熊小英, 黄芸培, 丁昌峰, 丁明军, 王鹏. 秧苗富锌对不同水稻品种镉积累及相关转运基因的影响[J]. 中国农业科学, 2022, 55(17): 3267-3277. DOI:10.3864/j.issn.0578-1752.2022.17.001 JIANG X T, HUANG G X, XIONG X Y, HUANG Y P, DING C F, DING M J, WANG P. Effects of seedlings enriched with zinc on cadmium accumulations and related transporter genes expressions in different rice cultivars[J]. Scientia Agriculture Sinica, 2022, 55(17): 3267-3277. DOI:10.3864/j.issn.0578-1752.2022.17.001 |
| [27] |
范锡麟, 杜强, 杨珍珍, 何艳冰, 王国梁, 王志龙. 水稻泛素连接酶基因OsRING6的克隆及表达分析[J]. 植物遗传资源学报, 2016, 17(2): 337-343. DOI:10.13430/j.cnki.jpgr.2016.02.019 FAN X L, DU Q, YANG Z Z, HE Y B, WANG G L, WANG Z L. Cloning and expression analysis of the rice ubiquitin ligase gene OsRING6[J]. Journal of Plant Genetic Resources, 2016, 17(2): 337-343. DOI:10.13430/j.cnki.jpgr.2016.02.019 |
| [28] |
WU Y F, WANG L L, ANSAH E O, PENG W M H, ZHANG W Y, LI P, AN G, XIONG F. The sucrose transport regulator OsDOF11 mediates cytokinin degradation during rice development[J]. Plant Physiology, 2022, 189(2): 1083-1094. DOI:10.1093/plphys/kiac104 |
| [29] |
刘晓月, 王文生, 傅彬英. 植物bHLH转录因子家族的功能研究进展[J]. 生物技术进展, 2011, 1(6): 391-397. DOI:10.3969/j.issn.2095-2341.2011.06.02 LIU X Y, WANG W S, FU B Y. Research progress of plant bHLH transcription factor family[J]. Current Biotechnology, 2011, 1(6): 391-397. DOI:10.3969/j.issn.2095-2341.2011.06.02 |
| [30] |
WANG K, LI M Q, ZHANG B, CHANG Y P, AN S H, ZHAO W L. Sugar starvation activates the OsSnRK1a-OsbHLH111/OsSGI1-OsTPP7 module to mediate growth inhibition of rice[J]. Plant Biotechnology Joumal, 2023, 21(10): 2033-2046. DOI:10.1111/pbi.14110 |
| [31] |
贾俊婷, 陈兵先, 吴柔贤, 戴彰言, 钟明生, 解昊, 刘双幸, 刘军. 水稻组蛋白去甲基化酶基因OsJMJ719的克隆及表达分析[J]. 广东农业科学, 2022, 49(11): 152-161. DOI:10.16768/j.issn.1004-874X.2022.11.016 JIA J T, CHEN B X, WU R X, DAI Z Y, ZHONG M S, XIE H, LIU S X, LIU J. Analysis on cloning and expression of histone demethylase gene OsJMJ719 in rice[J]. Guangdong Agricultural Sciences, 2022, 49(11): 152-161. DOI:10.16768/j.issn.1004-874X.2022.11.016 |
| [32] |
YAMAMURA C, MIZUTANI E, OKADA K, NAKAGAWA H, FUKUSHINA S, TANAKA A, MAEDA S, KAMAKURA T, YAMANE H, TAKATSUJI H, MORI M. Diterpenoid phytoalexin factor, a bHLH transcription factor, plays a central role in the biosynthesis of diterpenoid phytoalexins in rice[J]. The Plant Journal, 2015, 84(6): 1100-1113. DOI:10.1111/tpj.13065 |
| [33] |
YI K K, WU Z C, ZHOU J, DU L M, GUO L B, WU Y R, WU P. OsPTF1, a novel transcription factor involved in tolerance to phosphate starvation in rice[J]. Plant Physiology, 2005, 138(4): 2087-2096. DOI:10.1104/pp.105.063115 |
| [34] |
MAO K, DONG Q L, LI C, LIU C H, MA F W. Genome wide identification and characterization of apple bHLH transcription factors and expression analysis in response to drought and salt stress[J]. Frontiers in Plant Science, 2017, 8: 480. DOI:10.3389/fpls.2017.00480 |
| [35] |
KIRIBUCHI K, SUGIMORI M, TAKEDA M, OTANI T, OKADA K, ONODERA H, UGAKI M, TANAKA Y, TOMIYAMA-AKIMOTO C, YAMAGUCHI T, MINAMI E, SHIBUYA N, OMORI T, NISHIYAMA M, NOJIRI H, YAMANE H. RERJ1, a jasmonic acid-responsive gene from rice, encodes a basic helix-loop-helix protein[J]. Biochemical and Biophysical Research Communications, 2004, 325(3): 857-863. DOI:10.1016/j.bbrc.2004.10.126 |
| [36] |
KIRIBUCHI K, JIKUMARU Y, KAKU H, MINAMI E, HASEGAWA M, KODAMA O, SETO H, OKADA K, NOJIRI H, YAMANE H. Involvement of the basic Helix-Loop-Helix transcription factor RERJ1 in wounding and drought stress responses in rice plants[J]. Biochemical and Biophysical Research Communications, 2005, 69(5): 1042-1044. DOI:10.1271/bbb.69.1042 |
| [37] |
张子佳, 王迪, 傅彬英. 水稻转录因子bHLH家族基因响应环境胁迫表达谱分析[J]. 分子植物育种, 2008(3): 425-431. DOI:10.3969/j.issn.1672-416X.2008.03.003 ZHANG Z J, WANG D, FU B Y. Expression patterns of rice bHLH genes responsive to environmental stresses[J]. Molecular Plant Breeding, 2008(3): 425-431. DOI:10.3969/j.issn.1672-416X.2008.03.003 |
| [38] |
葛国平, 吴亮宇, 张少雄, 林金科. 茶树芽叶儿茶素含量与转录因子MYB、bHLH关联分析[J]. 广东农业科学, 2014, 41(9): 28-32. DOI:10.16768/j.issn.1004-874X.2014.09.034 GE G P, WU L Y, ZHANG S X, LIN J K. Correlation analysis between catechins content and transcription factor MYB, bHLH of bud and leaf in tea plant[J]. Guangdong Agricultural Sciences, 2014, 41(9): 28-32. DOI:10.16768/j.issn.1004-874X.2014.09.034 |
| [39] |
MENG F W, YANG C, CAO J D, CHEN H, PANG J H, ZHAO Q Q, WANG Z Y, FU Z Q, LIU J. A bHLH transcription activator regulates defense signaling by nucleo cytosolic trafficking in rice[J]. Journal of Integrative Plant Biology, 2020, 62(10): 1552-1573. DOI:10.1111/jipb.12922 |
| [40] |
DONG H J, ZHAO H, LI S L, HAN Z M, HU G, LIU C, YANG G Y, WANG G W, XIE W B, XING Y Z. Genome-wide association studies reveal that members of bHLH subfamily 16 share a conserved function in regulating flag leaf angle in rice (Oryza sativa)[J]. Plos Genetics, 2018, 14(4): e1007323. DOI:10.1371/journal.pgen.1007323 |
| [41] |
周扬, 王鹏, 李雨欣. 植物耐盐分子机制研究进展[J]. 广东农业科学, 2023, 50(10): 97-109. DOI:10.16768/j.issn.1004-874X.2023.10.011 ZHOU Y, WANG P, LI Y X. Research progress on molecular mechanism of plant salt tolerance[J]. Guangdong Agricultural Sciences, 2023, 50(10): 97-109. DOI:10.16768/j.issn.1004-874X.2023.10.011 |
| [42] |
WU H, YE H Y, YAO R F, ZHANG T, XIONG L T. OsJAZ9 acts as a transcriptional regulator in jasmonate signaling and modulates salt stress tolerance in rice[J]. Plant Science, 2015, 232: 1-12. DOI:10.1016/j.plantsci.2014.12.010 |
| [43] |
WANG Y J, ZHANG Z G, HE X J, ZHOU H L, WEN Y X, DAI J X, ZHANG J S, CHEN S Y. A rice transcription factor OsbHLH1 is involved in cold stress response[J]. Theoretical & Applied Genetics, 2003, 107(8): 1402-1409. DOI:10.1007/s00122-003-1378-x |
(责任编辑 陈丽娥)