 2023, Vol. 50
2023, Vol. 50文章信息
引用本文 |

基金项目
- 广东省乡村振兴战略专项资金种业振兴项目(2022NPY01004);广州市科技计划项目(202002020086);广州市支农资金项目(21102266)
作者简介
- 高银(1997—),男,在读硕士生,研究方向为蔬菜遗传育种,E-mail:gao1yin@qq.com.
通讯作者
- 乔燕春(1979—),女,博士,研究员,研究方向为蔬菜育种与栽培,E-mail:516986554@qq.com.
文章历史
- 收稿日期:2023-02-26




2. 广州市农业科学研究院,广东 广州 510308
2. Guangzhou Academy of Agricultural Sciences, Guangzhou 510308, China
【研究意义】种子大小与质量是重要的农艺性状,种子大小对出苗和幼苗生长均有一定的影响,同时决定了作物的产量[1]。一般来说,较大的种子生长出来的种苗比小种子生长出来的种苗更大[2-3]。在相同的生长环境中,大种和小种都能保证出苗率不受影响或影响较小,但对种苗后期的发育影响较大,尤其是在不利的环境中,大种子的优势更为明显,其相较于小种子来说生长更旺盛、产量更高,而小种子则能有效地分散和定植[4]。因此,探究种子性状对作物改良种质、提高产量以及解决生物能源问题具有重要作用。【前人研究进展】目前已在很多作物上开展了种子大小的遗传研究,王敏等[5]对黄瓜6个世代进行遗传分析,得到黄瓜种子宽度和长度的遗传均符合加性-显性-上位性多基因遗传模型,且以多基因加性效应为主;张可鑫等[6]得到甜瓜种子长度、宽度和百粒质量的遗传分别符合2对加性-显性-上位性主基因遗传模型以及1对加性-显性主基因遗传模型;王毅等[7]通过对冬瓜4个世代遗传分析,得出冬瓜种子长度的遗传符合2对完全显性主基因+加性-显性主基因遗传模型(E5),种子宽度的遗传符合2对加性-显性主基因+加性-显性多基因模型(E2);刘文睿等[8]通过对冬瓜6个世代种子千粒质量进行遗传分析,得出冬瓜千粒质量的遗传符合1对加性主基因+加性-显性多基因遗传模型;周延峰等[9]研究发现,西瓜种子长度的遗传符合D-3模型(1对完全显性主基因+加性-显性多基因遗传模型),种子宽度和百粒质量的遗传则均符合D-2模型(1对加性主基因+加性-显性多x基因模型);赵天波等[10]发现西葫芦种子长度和宽度的遗传分别符合B-1模型(2对加性-显性-上位性主基因遗传模型)和E-1模型(2对加性-显性-上位性主基因+加性-显性多基因混合遗传模型);邹一超等[11]研究发现,苦瓜种子长度的遗传符合加性-显性-上位性多基因遗传模型(C-0模型),种子宽度的遗传符合D-0模型。节瓜〔Benincasa hispida (Thunb.) Cogn. var. chiehqua How〕又名毛瓜,是葫芦科冬瓜属的一个变种,为藤蔓生草本植物,是广东省主要蔬菜作物,其产品除供应本地市场外还销往港澳地区,种植效益显著。【本研究切入点】相对于拟南芥、水稻等植物种子大小的研究,瓜类种子大小的研究仍处于初级阶段,节瓜作为我国华南地域性作物,关于种子大小的遗传研究还未见报道。【拟解决的关键问题】本研究以种子大小差异显著的节瓜高世代自交系为亲本,构建P1、P2、F1和F2 4个世代群体,并采用主基因+多基因混合模型对种子的长度、宽度和30粒种子质量进行遗传分析,揭示种子大小的遗传规律,以期为节瓜种子大小的基因定位、选育新品种并获得更大经济效益提供理论参考。
1 材料与方法 1.1 试验材料供试节瓜多世代稳定自交系材料均由广州市农业科学研究院提供。以大粒种子节瓜自交系J16为母本(P1)、小粒种子节瓜自交系18FJ5为父本(P2),通过杂交获得F1(图 1),再将F1自交获得F2群体。J16(P1)种子长度、宽度以及30粒种子质量分别为11.44(±0.27)mm、5.97(±0.14)mm、1.34(±0.1)g;18FJ5(P2)种子长度、宽度以及30粒种子质量分别为6.5(±0.24)mm、3.43(±0.15)mm、0.62(±0.07)g。

|
| 图 1 节瓜亲本及F1代种子大小比较 Fig. 1 Comparison of seed size between Chieh-qua parents and F1 generation |
1.2 性状测定
2021年7月,同时将4个世代的种子进行浸泡、催芽以及育苗,并定植于广州市农业科学研究院南沙试验基地。行距为0.8 m,株距为0.3 m,人工授粉前及授粉后对雌花和雄花进行隔离,避免其他物种干扰,且每株只留2个瓜,待节瓜表面布满白粉或毛绒褪掉时进行采摘取种。取母本20株、父本20株、F1代30株、F2代236株,每株采收的种子混合在一起并取10粒颗粒饱满的种子,利用游标卡尺分别对种子长度和宽度进行测量,随后再称量30粒饱满种子总质量(万分之一天平),用于种子大小和质量的遗传分析。
1.3 数据分析利用Excel、SPSS软件对节瓜种子大小进行统计。利用遗传模型分析软件包对种子长度、宽度以及质量进行遗传模型分析,得出极大似然函数值(Max-likelihood-values,MLV)和AIC值(Akaike’s information criterion)。根据AIC准则,选择3个AIC值最小的模型作为候选模型并进行适合性检验。最后,通过软件包计算出最优遗传模型的一、二阶遗传参数,并对主基因、多基因等遗传效应值进行分析。
2 结果与分析 2.1 节瓜4个世代群体种子大小表型分析对节瓜3个世代种子长度、宽度以及30粒种子质量进行统计分析,结果(表 1)表明,F1代种子长度、宽度以及质量整体偏向于大粒亲本P1。在F2代群体中,种子长度、宽度以及质量3个性状出现分离,并均呈双峰分布(图 2),表明三者均符合数量性状特征。
|

|
| 图 2 节瓜种子长度、宽度和30粒种子质量在F2群体中的频率分布 Fig. 2 Frequency distribution of seed length, width and mass of 30 seeds in F2 population of Chieh-qua |
2.2 节瓜种子大小遗传模型分析
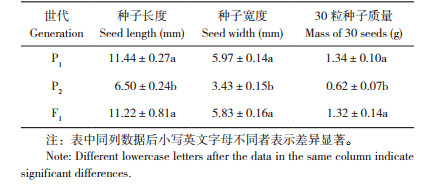
由表 2可知,节瓜种子长度的3个最小AIC值分别为850.3420、853.7899和856.4725,其对应的遗传模型分别是MX2-ADI-AD、MX1-AD-ADI和MX2-ADI-ADI,以这3组模型作为候选模型;种子宽度的3个最小AIC值分别为394.2935、395.1636和396.9915,对应的3个候选模型为MX1-AD-ADI、MX2-ADI-AD和MX2-A-AD;30粒种子质量的最小AIC值分别为89.4152、95.8995和90.9508,对应候选模型为MX1-AD-ADI、MX2-ADI-ADI和MX2-ADI-AD。
|
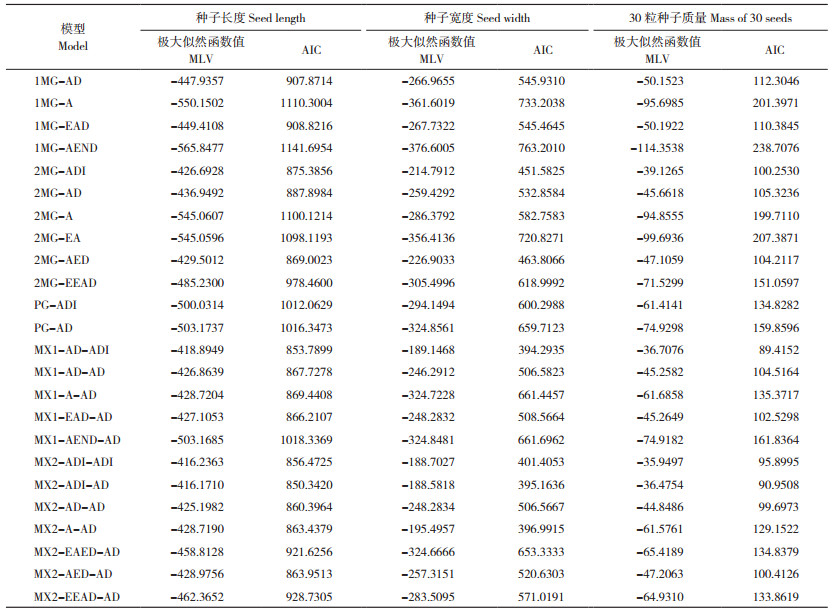
2.3 节瓜种子大小候选遗传模型的适合性检验
通过对节瓜以上3个性状的候选遗传模型进行适合性检验,选择AIC值最小或相对较小且统计为差异不显著的模型作为最优模型(表 3)。种子长度的3个候选遗传模型均在3个世代中达到显著,根据AIC准则,同样条件下MX2-ADI-AD模型的AIC值最小。因此,节瓜种子长度的最优遗传模型为MX2-ADI-AD,即2对加性-显性-上位性主基因+加性-显性多基因遗传模型。在种子宽度3个候选遗传模型中,MX2-ADI-AD模型的AIC值最小,且在各世代中均没有达到显著,因此节瓜种子宽度最优遗传模型为MX2-ADI-AD,即2对加性-显性-上位性主基因+加性-显性多基因遗传模型。30粒种子质量的候选遗传模型在各世代中均未达到显著,AIC值最小的MX1-AD-ADI模型则为最优遗传模型,即1对加性-显性主基因+加性-显性-上位性多基因遗传模型。
2.4 节瓜种子大小最适遗传模型的遗传参数估计
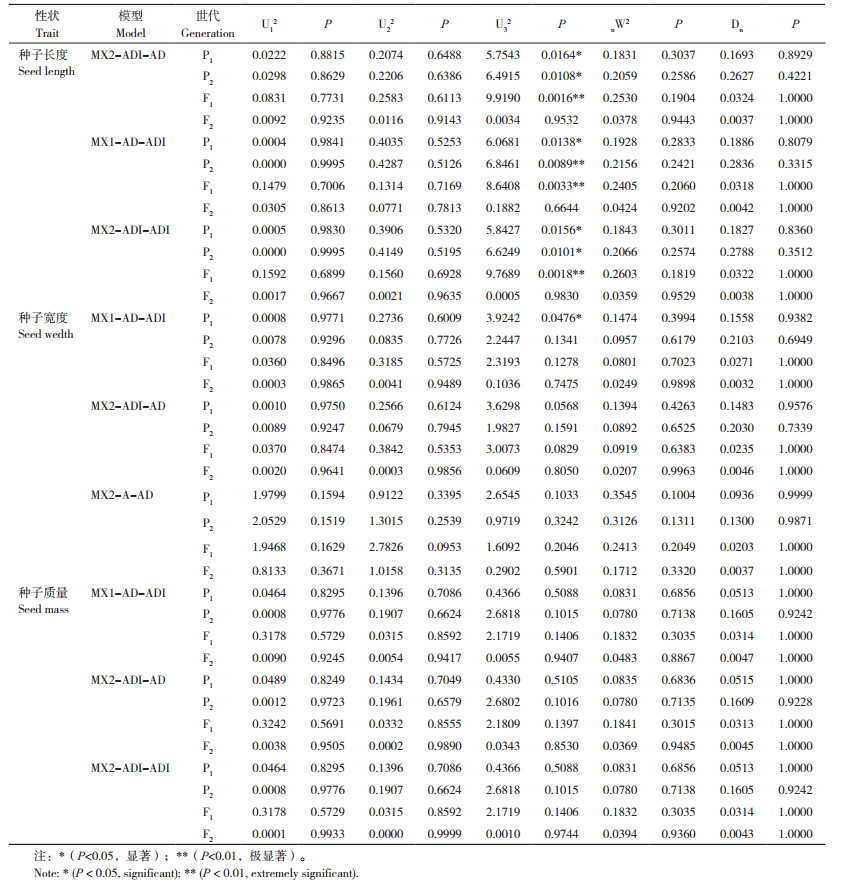
2.4.1 种子长度 由表 4可知,在一阶遗传参数中,第1对主基因的加性效应值(da)为2.0211,第2对主基因的加性效应值(db)为0.3035,2对主基因的加性效应值绝对值之和(da+db)为2.3246。2对主基因的显性效应值(ha和hb)分别为1.0577和-0.3169,显性效应值绝对值之和(ha+hb)为1.3746,表明主基因的加性效应在控制节瓜该性状中起主导作用。且第1对主基因的加性效应大于第2对主基因的加性效应(da>db),说明主基因以第1对的正向加性效应为主;0.1469和1.6188分别为多基因的加性效应值[d]和显性效应值[h],显性度大于1,说明起主导作用的是多基因的显性效应[h]。在主要互作效应中,加性×加性(i)、显性×显性(l)互作为正向,加性×显性(jab)和显性×加性(jba)互作均为负向。在二阶遗传参数中可以看出,控制节瓜种子长度的主基因遗传率(hmg2)为87.07%,而多基因遗传率(hPg2)为0,表明主基因对节瓜种子长度遗传的作用较大。
|
2.4.2 种子宽度 由节瓜种子宽度的一阶遗传参数可知,在控制该性状中起主导作用的是主基因的加性效应,且主基因以第1对的正向加性效应为主;加性效应值[d] 和显性效应值[h] 分别为-0.7634和1.5147,说明多基因的显性效应[h]起主导作用;节瓜种子宽度的主要互作效应情况与种子长度的遗传效应一致。在二级遗传参数中可以得到,控制节瓜种子宽度的主基因遗传率(hmg2)为90.79%,多基因遗传率(hPg2)为0,说明主基因对节瓜种子宽度遗传的作用较大。
2.4.3 种子质量 在30粒种子质量中,控制种子长度的主基因遗传率(hmg2)为79.96%,多基因遗传率(hPg2)为8.73%,同样说明了主基因在此性状中作用较大。
3 讨论种子是高等植物最主要的繁殖器官,是植物生命周期的起点和终点,种子对植物的生长发育至关重要,同时也是影响产量的重要因素。质量性状可以通过孟德尔遗传定律进行分析,主基因+多基因混合遗传模型分析方法解决了孟德尔遗传定律不能揭示的数量性状遗传规律,是数量性状遗传分析的经典方法[12],广泛应用于大豆[13]、水稻[14-16]、小麦[17-18]、玉米[19-20]、茄子[21-23]、花生[24-25]、烟草[26-27]、番茄[28]、辣椒[29-30]、瓠瓜[31]、丝瓜[32]等作物性状育种中,为遗传改良提供了重要信息。节瓜种子大小和质量的遗传分析鲜有报道,本研究对不同世代的节瓜种子长度、宽度及30粒种子质量进行测定,得出F1代的种子表型均接近大种亲本P1,在F2代群体中呈双峰分布。主基因+多基因遗传分析结果表明,节瓜种子长度和宽度的最适遗传模型均为MX2-ADI-AD,即2对加性-显性-上位性主基因+加性-显性多基因遗传模型,主基因遗传率分别为87.07%和90.79%、多基因遗传率均为0,其中节瓜种子宽度遗传模型与西葫芦种子宽度遗传模型一致[10],30粒种子质量的最适遗传模型为MX1-AD-ADI,即1对加性-显性主基因+加性-显性-上位性多基因遗传模型,主基因和多基因遗传率分别为79.96%和8.73%。通过遗传参数估计,可以发现主基因在这3个性状中作用较大,这与王敏等[5]研究结果一致。但除了主效基因,我们推测可能还存在其他微效基因对种子长度、宽度及质量起修饰作用。节瓜与冬瓜具有较近的亲缘关系,遗传背景非常窄[33],但本研究与王毅等[7]“冬瓜长度遗传符合2对完全显性主基因+加性-显性多基因遗传模型(E-5),宽度遗传符合2对加性-显性主基因+加性-显性多基因模型(E-2),千粒质量遗传符合1对加性主基因+加性-显性多基因遗传模型”的研究结果不一致。此外,节瓜种子长度、宽度以及30粒种子质量均呈双峰分布,表现为数量性状特征,与多数种子大小遗传研究的结果一致,但也有部分学者认为种子大小为质量性状,这可能是由于材料、种植方式或测量方法不同导致。今后仍需对不同遗传背景下节瓜种子大小的遗传模型进行分析。
4 结论本研究以大粒种子节瓜自交系J16(P1)和小粒种子节瓜自交系18FJ5(P2)为亲本进行杂交,结果发现F1群体的种子长度、宽度以及30粒质量均接近大粒亲本J16(P1),F2代出现分离,节瓜种子长度、宽度以及质量均呈双峰分布,表现为数量性状特征。对这些性状进行主基因+多基因遗传分析得出各性状最适遗传模型及遗传参数,发现主基因在种子长度、宽度以及质量3个性状中作用较大,其遗传率分别为87.07%、90.79%和79.96%;种子长度、宽度以加性效应为主,最佳遗传模型均为MX2-ADI-AD(2对加性-显性-上位性主基因+加性-显性多基因),30粒种子质量的最佳遗传模型为MX1-AD-ADI(1对加性-显性主基因+加性-显性-上位性多基因)。综上,可重视利用主基因,采用组合育种的途径对节瓜进行改良。
| [1] |
LI N, LI Y. Maternal control of seed size in plants[J]. Journal of Experimental Botany, 2015, 66(4): 1087-1097. DOI:10.1093/jxb/eru549 |
| [2] |
彭鸿嘉. 六种牧草种子大小和播种深度对出苗的影响[J]. 草业科学, 2001(6): 30-35. DOI:10.3969/j.issn.1001-0629.2001.06.008 PENG H J. Effects of seed size and seedling depth on emergence of six perennial grasses[J]. Pratacultural Science, 2001(6): 30-35. DOI:10.3969/j.issn.1001-0629.2001.06.008 |
| [3] |
向长萍, 史雪梅, 张亚. 苦瓜种子大小对种子质量及产量的影响[J]. 长江蔬菜, 2002(2): 35-36. DOI:10.3865/j.issn.1001-3547.2002.02.024 XIANG C P, SHI X M, ZHANG Y. Effect of bitter gourd seed size on seed quality and yield[J]. Journal of Chang jiang Vegetables, 2002(2): 35-36. DOI:10.3865/j.issn.1001-3547.2002.02.024 |
| [4] |
WESTOBY M, FALSTER D S, MOLES A T, VESK P A, WRIGHT I J. Plant ecological strategies: Some leading dimensions of variation between species[J]. Annual Review of Ecology and Systematics, 2002, 33. DOI:10.1146/annurev.ecolsys.33.010802.150452 |
| [5] |
王敏, 苗晗, 张圣平, 刘书林, 董邵云, 王烨, 顾兴芳. 黄瓜种子大小遗传分析与QTL定位[J]. 园艺学报, 2014, 41(1): 63-72. DOI:10.16420/j.issn.0513-353x.2014.01.010.j.issn.0513-353x.2014.01.010 WANG M, MIAO H, ZHANG S P, LIU S L, DONG S Y, WANG Y, GU X F. Inheritance analysis and QTL mapping of cucumber seed size[J]. Acta Horticulturae Sinica, 2014, 41(1): 63-72. DOI:10.16420/j.issn.0513-353x.2014.01.010.j.issn.0513-353x.2014.01.010 |
| [6] |
张可鑫, 戴冬洋, 王浩男, 蔚明月, 盛云燕. 甜瓜种子相关性状遗传规律与QTL分析[J]. 浙江农业学报, 2018, 30(9): 1496-1503. ZHANG K X, DAI D Y, WANG H N, WEI M Y, SHENG Y Y. Genetic and QTL analysis of seed traits in melon (Cucumis melon L.)[J]. Acta Agriculturae Zhejiangensis, 2018, 30(9): 1496-1503. |
| [7] |
王毅, 谢大森, 江彪, 王敏, 彭庆务, 何晓明, 林毓娥, 闫晋强, 梁肇均, 陈林, 刘文睿. 冬瓜种子大小的遗传分析及QTL定位// 中国园艺学会. 中国园艺学会2019年学术年会暨成立90周年纪念大会论文摘要集[C]. 2019: 193. DOI: 10.26914/c.cnkihy.2019.000824. WANG Y, XIE D S, JIANG B, WANG M, PENG Q W, HE X M, LIN Y E, YAN J Q, LIANG Z J, CHEN L, LIU W R. Genetic analysis and QTL mapping of seed size of wax gourd// Chinese Horticultural Society. Abstracts of the 2019 Academic Annual Conference and the 90th Anniversary of the Establishment of the Chinese Horticultural Society [C]. 2019: 193. DOI: 10.26914/c.cnkihy.2019.000824. |
| [8] |
刘文睿, 江彪, 彭庆务, 赵芹, 谢大森. 冬瓜种子千粒重主基因+多基因混合遗传分析[J]. 热带作物学报, 2012, 33(6): 993-996. DOI:10.3969/j.issn.1000-2561.2012.06.005 LIU W R, JIANG B, PENG Q W, ZHAO Q, XIE D S. Analysis of 1000-seed weight by mixed major and polygene inheritance model in Benincasa hispida cogn.[J]. Chinese Journal of Tropical Crops, 2012, 33(06): 993-996. DOI:10.3969/j.issn.1000-2561.2012.06.005 |
| [9] |
周延峰. 西瓜主要品质及种子性状遗传分析[D]. 北京: 中国农业科学院, 2014. ZHOU Y F. Genetic analysis of main quality and seed traits of watermelon[D]. Beijing: Chinese Academy of Agricultural Sciences, 2014. |
| [10] |
赵天波. 西葫芦种子形状遗传分析及制种技术优化[D]. 泰安: 山东农业大学, 2016. ZHAO T B. Genetic analysis of seed shape and optimization of seed production technology of summer squash (Cucurbitta pepo L. )[D]. Taian: Shandong Agricultural University, 2016. |
| [11] |
邹一超, 钟建, 胡芳, 董骥驰, 胡开林, 程蛟文. 苦瓜种子大小和单粒质量的遗传分析[J]. 中国蔬菜, 2020(4): 61-66. DOI:10.19928/j.cnki.1000-6346.2020.04.013 ZOU Y C, ZHONG J, HU F, DONG J C, HU K L, CHENG J W. Genetic analysis on seed size and single seed weight in bitter gourd (Momerdica charantia L.)[J]. China Vegetables, 2020(4): 61-66. DOI:10.19928/j.cnki.1000-6346.2020.04.013 |
| [12] |
盖钧镒, 章元明. 植物数量性状遗传体系检测的试验方法讨论// 中国作物学会. 21世纪作物科技与生产发展学术讨论会论文集[C]. 北京: 中国农业科学技术出版社, 2002: 36-42. GAI J Y, ZHANG Y M. Experiments methods for the detection of QTL system in planst// The Crop Science Society of China. Proceedings of the Symposium on Crop Science and Technology and Production Development in the 21st Century[C]. Beijing: China Agriculural Science and Technology Press, 2002: 36-42. |
| [13] |
张玉梅, 蓝新隆, 滕振勇, 陆佩兰, 林国强, 胡润芳. 大豆鲜籽粒可溶性糖含量的主基因+多基因遗传分析[J]. 大豆科学, 2022, 41(5): 520-525. DOI:10.11861/j.issn.1000-9841.2022.05.0520 ZHANG Y M, LAN X L, TENG Z Y, LU P L, LIN G Q, HU R F. Genetic analysis of fresh seeds soluble sugar content in soybean based on a mixed model of major-genes plus polygenes[J]. Soybean Science, 2022, 41(5): 520-525. DOI:10.11861/j.issn.1000-9841.2022.05.0520 |
| [14] |
陈志彬, 陈小荣. 水稻茎秆粗度主基因+多基因混合遗传分析[J]. 杂交水稻, 2010, 25(S1): 108-111. DOI:10.16267/j.cnki.1005-3956.2010.s1.081 CHEN Z B, CHEN X R. Analysis on major genes plus polygenes mixed inheritance of culm thickness trait in rice[J]. Hybrid Rice, 2010, 25(S1): 108-111. DOI:10.16267/j.cnki.1005-3956.2010.s1.081 |
| [15] |
陈小荣, 陈志彬, 贺浩华, 朱昌兰, 彭小松, 贺晓鹏, 傅军如, 欧阳林娟. 水稻单株有效穗数主基因+多基因混合遗传分析[J]. 生物数学学报, 2011, 26(3): 555-562. CHEN X R, CHEN Z B, HE H H, ZHU C L, PENG X S, HE X P, FU J R, OUYANG L J. Genetic analysis of panicles per plant in rice by the major genes plus polygenes mixed inheritance model[J]. Journal of Biomathematics, 2011, 26(3): 555-562. |
| [16] |
江建华, 张武汉, 党小景, 荣慧, 叶琴, 胡长敏, 张瑛, 何强, 王德正. 水稻核不育系柱头性状的主基因+多基因遗传分析[J]. 作物学报, 2021, 47(7): 1215-1227. DOI:10.3724/SP.J.1006.2021.02057 JIANG J H, ZHANG W H, DANG X J, RONG H, YE Q, HU C M, ZHANG Y, HE Q, WANG D Z. Genetic analysis of stigma traits with genic male sterile line by mixture model of major gene plus polygene in rice (Oryza sativa L.)[J]. Acta Agronomica Sinica, 2021, 47(7): 1215-1227. DOI:10.3724/SP.J.1006.2021.02057 |
| [17] |
解松峰, 吉万全, 张耀元, 张俊杰, 胡卫国, 李俊, 王长有, 张宏, 陈春环. 小麦重要产量性状的主基因+多基因混合遗传分析[J]. 作物学报, 2020, 46(3): 365-384. DOI:10.3724/SP.J.1006.2020.91044.SP.J.1006.2020.91044 XIE S F, JI W Q, ZHANG Y Y, ZHANG J J, HU W G, LI J, WANG C Y, ZHANG H, CHEN C H. Genetic effects of important yield traits analysed by mixture model of major gene plus polygene in wheat[J]. Acta Agronomica Sinica, 2020, 46(3): 365-384. DOI:10.3724/SP.J.1006.2020.91044.SP.J.1006.2020.91044 |
| [18] |
毕晓静, 史秀秀, 马守才, 韩芳, 亓佳佳, 李清峰, 王志军, 张改生, 牛娜. 小麦农艺性状的主基因+多基因遗传分析[J]. 麦类作物学报, 2013, 33(4): 630-634. DOI:10.7606/j.issn.1009-1041.2013.04.002 BI X J, SHI X X, MA S C, HAN F, QI J J, LI Q F, WANG Z J, ZHANG G S, NIU N. Genetic analysis of agronomic traits related to yield based on major gene plus polygene model in wheat[J]. Journal of Triticeae Crops, 2013, 33(4): 630-634. DOI:10.7606/j.issn.1009-1041.2013.04.002 |
| [19] |
胡洋, 崔学宇, 郑雷雷, 杨伟光. 玉米穗部性状遗传分析[J]. 广东农业科学, 2014, 41(7): 12-15. DOI:10.16768/j.issn.1004-874X.2014.07.019 HU Y, CUI X Y, ZHENG L L, YANG W G. Genetic analysis of maize ear characters[J]. Guangdong Agricultural Sciences, 2014, 41(7): 12-15. DOI:10.16768/j.issn.1004-874X.2014.07.019 |
| [20] |
蒋锋, 梁日朗, 闫艳, 梁泽恩, 黄正刚, 刘鹏飞. 甜玉米果皮厚度遗传分析及QTL定位[J]. 广东农业科学, 2022, 49(8): 21-28. DOI:10.16768/j.issn.1004-874X.2022.08.003 JIANG F, LIANG R L, YAN Y, LIANG Z E, HUANG Z G, LIU P F. Genetic analysis and QTL Mapping of pericarp thickness in sweet corn[J]. Guangdong Agricultural Sciences, 2022, 49(8): 21-28. DOI:10.16768/j.issn.1004-874X.2022.08.003 |
| [21] |
乐素菊, 汪文毅, 邵光金, 汪国平. 茄子果形性状的主基因+多基因混合模型遗传分析[J]. 华南农业大学学报, 2011, 32(3): 27-31. DOI:10.3969/j.issn.1001-411X.2011.03.007 LE S J, WANG W Y, SHAO G J, WANG G P. Analysis on mixed major gene and polygene inheritance of fruit morphological traits in eggplant[J]. Journal of South China Agricultural University, 2011, 32(3): 27-31. DOI:10.3969/j.issn.1001-411X.2011.03.007 |
| [22] |
王倩, 崔庭源, 孙国胜, 马志虎, 张长青. 茄子叶色性状的主基因+多基因遗传模型分析[J]. 金陵科技学院学报, 2020, 36(2): 88-92. DOI:10.16515/j.cnki.32-1722/n.2020.02.018 WANG Q, CUI T Y, SUN G S, MA Z H, ZHANG C Q. Genetic analysis of leaf color in eggplant with major gene plus polygene model[J]. Journal of Jinling Institute of Technology, 2020, 36(2): 88-92. DOI:10.16515/j.cnki.32-1722/n.2020.02.018 |
| [23] |
张永楠, 黄诗诗, 胡胜平, 王鹏, 钟川, 阳燕娟, 于文进. 茄子果实形状的主基因+多基因遗传分析[J/OL]. 分子植物育种, 2023: 1-14. http://kns.cnki.net/kcms/detail/46.1068.S.20210611.1644.024.html. ZHANG Y N, HUANG S S, HU S P, WANG P, ZHONG C, YANG Y J, YU W J. Genetic analysis of eggplant fruit shape by mixture model of major genes and polygenes[J/OL]. Molecular Plant Breeding, 2023: 1-14. http://kns.cnki.net/kcms/detail/46.1068.S.20210611.1644.024.html. |
| [24] |
张晓霞, 杨会, 张秀荣, 骆璐, 吕玉英, 张昆, 刘风珍, 万勇善. 花生子仁长宽及单仁重的遗传分析[J]. 山东农业科学, 2019, 51(9): 73-78, 86. DOI:10.14083/j.issn.1001-4942.2019.09.011 ZHANG X X, YANG H, ZHANG X R, LUO L, LYU Y Y, ZHANG K, LIU F Z, WAN Y S. Genetic analysis of kernel length, width and single kernel weight in peanut (Arachis hypogaea L.)[J]. Shandong Agricultural Sciences, 2019, 51(9): 73-78, 86. DOI:10.14083/j.issn.1001-4942.2019.09.011 |
| [25] |
牟大林, 韩笑, 李雪莹, 姚丹, 杨松楠, 曲艺伟, 梁嘉宁, 张君. 花生主要品质性状的主基因+多基因遗传分析[J]. 花生学报, 2021, 50(1): 41-44, 49. DOI:10.14001/j.issn.1002-4093.2021.01.005 MOU D L, HAN X, LI X Y, YAO D, YANG S N, QU Y W, LIANG J N, ZHANG J. Genetic analysis of major gene plus polygene on main quality traits in peanut[J]. Journal of Peanut Science, 2021, 50(1): 41-44, 49. DOI:10.14001/j.issn.1002-4093.2021.01.005 |
| [26] |
张兴伟, 王志德, 刘艳华, 任民, 杨洋. 植物数量性状"主基因+多基因"混合遗传模型及其在烟草上的应用[J]. 中国烟草学报, 2013, 19(3): 41-44. DOI:10.3969/j.issn.1004-5708.2013.03.008 ZHANG X W, WANG Z D, LIU Y H, REN M, YANG Y. Major gene plus polygene mixed genetic model in quantitative characters and its application in tobacco[J]. Acta Tabacaria Sinica, 2013, 19(3): 41-44. DOI:10.3969/j.issn.1004-5708.2013.03.008 |
| [27] |
吴兴富, 焦芳婵, 陈学军, 方敦煌, 张谊寒, 高玉龙, 童治军. 烟草主要农艺性状的主基因+多基因遗传分析[J]. 分子植物育种, 2021, 19(19): 6438-6447. DOI:10.13271/j.mpb.019.006438 WU X F, JIAO F C, CHEN X J, FANG D H, ZHANG Y H, GAO Y L, TONG Z J. Genetic analysis of main agronomic traits in tobacco (Nicot iana tabacum L.) by mixture model of major genes and polygenes[J]. Molecular Plant Breeding, 2021, 19(19): 6438-6447. DOI:10.13271/j.mpb.019.006438 |
| [28] |
马雅琳, 梁燕. 番茄长花柱性状遗传规律与QTL分析[J]. 园艺学报, 2020, 47(2): 264-274. DOI:10.16420/j.issn.0513-353x.2019-0458 MA Y L, LIANG Y. Inheritance and QTL analysis of long-style trait in tomato[J]. Acta Horticulturae Sinica, 2020, 47(2): 264-274. DOI:10.16420/j.issn.0513-353x.2019-0458 |
| [29] |
陈学军, 方荣, 周坤华, 缪南生, 黄长林. 辣椒果实性状主基因+多基因遗传分析[J]. 西北植物学报, 2012, 32(2): 246-251. DOI:10.3969/j.issn.1000-4025.2012.02.005 CHEN X J, FANG R, ZHOU K H, MIAO N S, HUANG C L. Major gene plus polygene inheritance analysis of fruit traits in pepper[J]. Acta Botanica Boreali-Occidentalia Sinica, 2012, 32(2): 246-251. DOI:10.3969/j.issn.1000-4025.2012.02.005 |
| [30] |
王俊涛, 冯鹏龙, 王亚艺, 李全辉. 辣椒单株结果数性状的主基因+多基因遗传分析[J]. 西北农业学报, 2022, 31(11): 1443-1450. DOI:10.7606/j.issn.1004-1389.2022.11.007 WANG J T, FENG P L, WANG Y Y, LI Q H. Genetic analysison pepper (Capsicum annuum L.) fruit number per plant by mixture model of major gene plus polygene[J]. Acta Agriculturae Boreali-Occidentali Sinica, 2022, 31(11): 1443-1450. DOI:10.7606/j.issn.1004-1389.2022.11.007 |
| [31] |
胡胜平, 霍雨来, 黄诗诗, 张永楠, 沈慕洁, 赵荣茺, 于文进, 阳燕娟. 瓠瓜果实形状的主基因+多基因遗传分析[J]. 分子植物育种, 2022, 20(13): 4407-4415. DOI:10.13271/j.mpb.020.004407 HU S P, HUO Y L, HUANG S S, ZHANG Y N, SHEN M J, ZHAO R C, YU W J, YANG Y J. Genetic analysis of bottle gourd fruit shape by mixture model of major genes and polygenes[J]. Molecular Plant Breeding, 2022, 20(13): 4407-4415. DOI:10.13271/j.mpb.020.004407 |
| [32] |
崔竣杰, 程蛟文, 谭澍, 李卫鹏, 汪国平, 胡开林. 丝瓜果长及果柄长的遗传规律分析[J]. 广东农业科学, 2014, 41(8): 52-56. DOI:10.16768/j.issn.1004-874X.2014.08.032 CUI J J, CHENG J W, TAN S, LI W P, WANG G P, HU K L. Genetic analysis of fruit length and fruit stalk length in luffa[J]. Guangdong Agricultural Sciences, 2014, 41(8): 52-56. DOI:10.16768/j.issn.1004-874X.2014.08.032 |
| [33] |
乔燕春, 林锦英, 谢伟平, 谢丽芳, 李莲芳. 节瓜、冬瓜重要性状多样性及亲缘关系的SRAP分析[J]. 植物遗传资源学报, 2014, 15(5): 1150-1155. DOI:10.13430/j.cnki.jpgr.2014.05.034 QIAO Y C, LIN J Y, XIE W P, XIE L F, LI L F. Genetic relationship of Chieh-qua and Wax-gourd on the morphological phenotypes and SRAP markers[J]. Journal of Plant Genetic Resources, 2014, 15(5): 1150-1155. DOI:10.13430/j.cnki.jpgr.2014.05.034 |
(责任编辑 马春敏)